(一)被子植物叶的一般结构

一般被子植物的叶片有上下面的区别,上面(即腹面或近轴面)深绿色,下面(即背面或远轴面)淡绿色,这种叶是由于叶片在枝上的着生取横向的位置,近乎和枝的长轴垂直或与地面平行,叶片的两面受光的情况不同,因而两面的内部结构也不同,即组成叶肉的组织有较大的分化,形成栅栏组织和海绵组织,这种叶称为异面叶(dorsi-ventralleaf或bifacialleaf)。有些植物的叶取近乎直立的位置,近乎和枝的长轴平行或与地面垂直,叶片两面的受光情况差异不大,因而叶片两面的内部结构也就相似,即组成叶肉的组织分化不大(图3-91),这种叶称为等面叶(isobilateral leaf)。有些植物的叶上下面都同样地具有栅栏组织,中间夹着海绵组织,也称等面叶。不论异面叶还是等面叶,就叶片来讲,都有三种基本结构,即表皮、叶肉和叶脉(图3-92)。表皮是包在叶的最外层,有保护作用;叶肉是在表皮的内方,有制造和贮藏养料的作用;叶脉是埋在叶肉中的维管组织,有输导和支持的作用。叶片的形态和结构尽管有多种多样,但是这三种基本结构总是存在的,只不过是形状、排列和数量的变化而已。
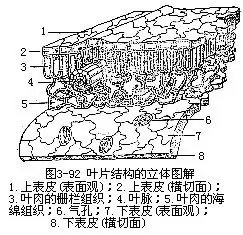

1.表皮表皮包被着整个叶片,有上下表皮之分。表皮通常由一层生活的细胞组成,但也有多层细胞组成的,称为复表皮(multiple pyidermis),如夹竹桃和印度橡胶树(Ficuselastica)叶的表皮(图3-93)。叶的表皮细胞在平皮切面(与叶片表面成平行的切面)上看,一般是形状规则或不规则的扁平细胞。不少双子叶植物叶表皮细胞的径向壁往往凹凸不平,犬牙交错地彼此镶嵌着,成为一层紧密而结合牢固的组织。在横切面上,表皮细胞的外形较规则,呈长方形或方形,外壁较厚,常具角质层。角质层的厚度因植物种类和所处环境而异。角质层是由表皮细胞内原生质体分泌所形成,通过质膜,沉积在表皮细胞的外壁上。多数植物叶的角质层外,往往还有一层不同厚度的蜡质层。近年来通过透射电子显微镜对表皮超微切片的观察,对角质层有了更好的理解(参阅图1-40,C)。角质层的存在,起着保护作用,可以控制水分蒸腾,加固机械性能,防止病菌侵入,对药液也有着不同程度的吸收能力。因此,角质层的厚壁,可作为作物优良品种选育时的根据之一。一般植物叶的表皮细胞不具叶绿体。表皮毛的有无和毛的类型也因植物的种类而异。
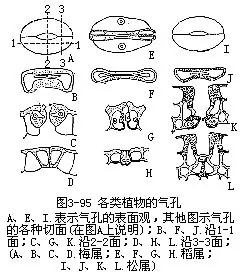
叶的表皮具有较多的气孔,这是和叶的功能有密切联系的一种绪构,它既是与外界进行气体交换的门户,又是水气蒸腾的通道,根外施肥和喷洒农药由此进入,因而也是水液的入口。各种植物的气孔数目、形态结构和分布是不同的(图3-94,图3-95)。
这里以双子叶植物的气孔为例,就气孔与相邻细胞的关系,即相邻细胞中有无副卫细胞(subsidiary cell),以及它的数目、大小与排列等为依据可分为四个主要类型:
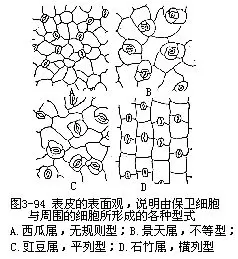
(1)无规则型(anomocytictype),也称毛茛科型(ranunculaceoustype),几个与表皮细胞大小、形状相同的细胞,无规则地围绕着保卫细胞,也就是无副卫细胞。所谓副卫细胞就是邻近保卫细胞的与一般表皮细胞大小形状有区别的细胞。本类型通常见于毛茛科(如毛茛属)、葫芦科(如西瓜属)以及其他科、属中(图3-94,A)。
(2)不等型(anisocytictype),也称十字花科型(cruciferoustype),有三个大小不同的副卫细胞围绕着保卫细胞,其中一个显著地较其他二者为小。本类型通常见于十字花科(如芸苔属)、景天科(如景天属)以及其他科、属中(图3-94,B)。
(3)平列型(paracytictype),也称茜草科型(rubiaceoustype),在每一保卫细胞侧面伴随着一个或几个副卫细胞,它们的长轴与气孔的长轴平行。本类型通常见于茜草科(如茜草属)、蝶形花科(如菜豆属)以及其他科、属中(图3-94,C)。
(4)横列型(diacytictype),也称石竹科型(caryophyllaceoustype),每一气孔由二副卫细胞围绕着,它们的共同壁与气孔的长轴形成直角。本类型通常见于石竹科(如石竹属)、爵床科(爵床属)以及其他科、属中(图3-94,D)。
气孔是由保卫细胞和它们间的孔口共同组成的。如果副卫细胞存在,副卫细胞及气孔又共同组成气孔器(stomatalapparatus)或称气孔复合体(stomatalcomplex)。气孔和气孔器的各种类型在分类学的鉴定上是有一定的价值。不同学者对气孔类型的制订,存在着不同的看法。各种植物的气孔和气孔器由于形状和结构不同,在表面观和各切面观上存在着显著的差异(图3-95)。例如单子叶植物稻属的气孔由于副卫细胞和哑铃状保卫细胞的存在,在表面观和切面观上都另具特点。裸子植物松属针叶的气孔,木质化较大和气孔下陷,也显出在切面观上与其他植物不同。
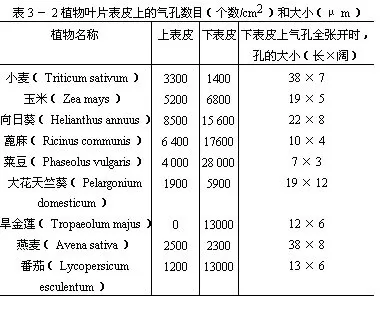
表3-2是几种气孔在叶片上、下表皮上分布的情况。
从表3-2可以看出气孔的数目和分布情况,在各个植物的叶中是不同的。此外,植物体上部叶的气孔较下部的多,叶尖端和中脉部分的气孔较叶基部和叶缘的多。有些植物如向日葵、蓖麻、玉米、小麦等叶的上、下表皮都有气孔,而下表皮一般较多。但也有些植物,气孔却只限于下表皮(如旱金莲、苹果)或限于上表皮(如莲、睡莲),还有些植物的气孔却只限于下表皮的局部区域,如夹竹桃叶的气孔,仅生在凹陷的气孔窝部分。在不同的外界环境中,同一种植物的叶气孔的数目,也有差异,一般阳光充足处较多,阴湿处较少。沉水的叶一般没有气孔(如眼子菜)。夹竹桃和眼子菜的叶,将在以后“叶的生态类型”中再作叙述。
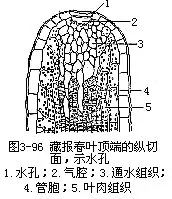
在叶尖或叶缘的表皮上,还有一种类似气孔的结构,保卫细胞长期开张,称为水孔(waterpore),是气孔的变形(图3-96)。

2.叶肉 叶肉是上、下表皮之间的绿色组织的总称,是叶的主要部分。通常由薄壁细胞组成,内含丰富的叶绿体。一般异面叶中,近上表皮部位的绿色组织排列整齐(图3-97),细胞呈长柱形,细胞长轴和叶表面相垂直,呈栅栏状,称为栅栏组织。其层数,因植物种类而异。栅栏组织的下方,即近下表皮部分的绿色组织,形状不规则,排列不整齐,疏松和具较多间隙,作海绵状,称为海绵组织。它和栅栏组织对比,排列较疏松,间隙较多,细胞内含叶绿体也较少。叶片上面绿色较深,下面较淡,就是由于两种组织内叶绿体的含量不同所致。光合作用主要是在叶肉中进行。
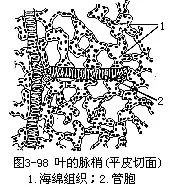
3.叶脉 前面已经讲过叶片上叶脉的分布状态,现在再讲叶脉在叶片中的内部结构。叶脉也就是叶内的维管束,它的内部结构,因叶脉的大小而不同。例如粗大的中脉(即中肋),它的内部结构是由维管束和伴随的机械组织组合而成。叶片中的维管束是通过叶脉而与茎中的维管束相连接。在茎中,维管束的木质部在内方,韧皮部在外方,进入叶片后,木质部却在上方(近轴面),而韧皮部在下方(远轴面),这是由于维管束从茎中向外方,侧向地进入叶中必然的结果。维管束外,还有由薄壁组织组成的维管束鞘包裹着。有些植物的叶具束鞘延伸也称维管束鞘延伸(bundlesheathextension),即组成束鞘的一些细胞延伸至上、下表皮,或一面的表皮上。在中脉和较大的叶脉中,维管束常相当发达,并有形成层,不过形成层的活动有限和活动期间较短,因而产生的次生组织不多。大的叶脉在维管束的上下方常有相当量的机械组织,直接和表皮相连接,机械组织在下方更为发达,因此,叶片的下面常有显著的凸出。在叶中叶脉越分越细,结构也愈来愈简化。就简化的趋向程度而言,一般首先是形成层的消失;其次,是机械组织的逐渐减少,以至完全不存在;再次,是木质部和韧皮部的结构简化。中等的脉,一般纯为初生结构,机械组织或有或无,即使存在,也不及大脉中的那样发达。叶片中最后的叶脉分枝,终止于叶肉组织内,往往成为游离的脉梢,结构异常简单,木质部,仅为一个螺纹管胞,而韧皮部仅有短狭的筛管分子和增大的伴胞,甚至有时只有木质部分子存在(图3-98)。

以上所讲的表皮、叶肉和叶脉等三种基本结构,在叶片中是普遍存在的,但是由于叶肉组织分化和发达的程度,栅栏组织的有无、层数和分布情况,海绵组织的有无和排列的疏松程度,气孔的类型和分布,以及表皮毛的有无和类型,都使叶片的结构在不同植物和不同生境中,有不少的变化。
从上述的叶片结构,可以看出,叶肉是叶的主要结构,是叶的生理功能主要进行的场所。表皮包被在外,起保护作用,使叶肉得以顺利地进行工作。叶脉分布于内,一方面,源源不绝地供应叶肉组织所需的水分和盐类,同时运输出光合的产物;另一方面,又支撑着叶面,使叶片舒展在大气中,承受光照。三种基本结构的合理组合和有机联系,也就保证叶片生理功能的顺利进行,这也表明叶片的形态、结构是完全适应它的生理功能的。
以上讲的是叶片的结构,现在再讲一下叶柄的结构(图3-99)。叶柄的结构比叶片要简单些,它和茎的结构有些相似,是由表皮、基本组织和维管组织组成的。在一般情况下,叶柄在横切面上通常呈半月形、圆形、三角形等。最外层为表皮;表皮内为基本组织,基本组织中近外方的部分往往有多层厚角组织,内方为薄壁组织;基本组织以内为维管束,数目和大小不定,排列成弧形、环形、平列形。维管束的结构和幼茎中的维管束相似,但木质部在上方(近轴面),韧皮部在下方(远轴面)。每一维管束外,常有厚壁的细胞包围。双子叶植物的叶柄中,木质部与韧皮部之间往往有一层形成层,但形成层只有短期的活动。在叶柄中由于维管束的分离和联合,使维管束的数目和排列变化极大,造成它的结构复杂化。
有托叶的叶,如果托叶外形是叶状的,它的结构一般和叶片的结构大致相似。
(二)单子叶植物叶的特点
单子叶植物的叶,就外形讲,有多种多样,如线形(稻、麦)、管形(葱)、剑形(鸢尾)、卵形(玉簪)、披针形(鸭跖草)等。叶脉多数为平行脉,少数为网状脉(薯蓣、菝葜等)。现以禾本科植物的叶为例,就内部结构加以说明。
前面讲过,禾本科植物叶的外形是叶片狭长,叶鞘包在茎外,在叶鞘与叶片连接处,有叶舌和叶耳。禾本科植物的叶片和一般叶一样,具有表皮、叶肉和叶脉三种基本结构(图3-100)。
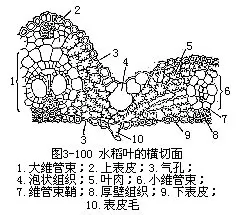
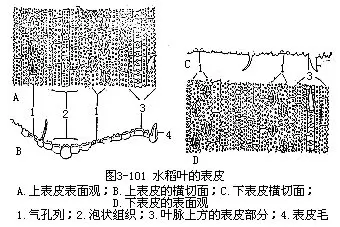
1.表皮表皮细胞的形状比较规则(图3-101),排列成行,常包括长、短两种类型的细胞。长细胞作长方柱形,长径与叶的纵长轴方向一致,横切面近乎方形,细胞壁不仅角质化,并且充满硅质,这是禾本科植物叶的特征;短细胞又分为硅质细胞和栓质细胞两种。硅质细胞常为单个的硅质体所充满,禾本科植物的叶,往往质地坚硬,易戳破手指就是由于含有硅质;栓质细胞是一种细胞壁栓质化的细胞,常含有有机物质。在表皮上,往往是一个长细胞和两个短细胞(即一个硅质细胞和一个栓质细胞)交互排列,有时也可见多个短细胞聚集在一起。长细胞与短细胞的形状、数目和相对位置,因植物种类而不同。禾本科植物叶的上、下表皮上,都有气孔,成纵行排列,与一般植物不同(图3-101)。保卫细胞呈哑铃形,中部狭窄,具厚壁,两端膨大,成球状,具薄壁。气孔的开阔是两端球状部分胀缩变化的结果。当两端球状部分膨胀时,气孔开放,反之,收缩时气孔关闭。保卫细胞的外侧各有一个副卫细胞,它和一般表皮细胞形状不同,有时甚至是内含物不同的细胞。这些副卫细胞骤看起来,仿佛是气孔的一部分,但实际上,它们都是由气孔侧面的表皮细胞所衍生。气孔的分布和叶脉相平行。气孔的数目和分布,因植物种类而异。同一植株的不同叶上,或同一叶上的不同部分,气孔的数目也有差别。上、下表皮上,气孔的数目近乎相等。在上表皮的不少地方,还有一些特殊的大型含水细胞,有较大的液泡,无叶绿素,或有少量的叶绿素,径向细胞壁薄,外壁较厚,称为泡状细胞(bulliformcell,图3-102)。泡状细胞通常位于两个维管束之间的部位,在叶上排列成若干纵列,列数因植物种类而不同。在横切面上,泡状细胞的排列略呈扇形。过去一般认为泡状细胞和叶片的伸展卷缩有关,即水分不足时,泡状细胞失水较快,细胞外壁向内收缩,引起整个叶片向上卷缩成筒,以减少蒸腾;水分充足时,泡状细胞膨胀,叶片伸展,因此,泡状细胞也称为运动细胞(motorcell)。但是有些试验表明,叶片的伸展、卷缩,最重要的是泡状细胞以外的其他组织如表皮、叶肉等的收缩。
2.叶肉叶肉组织比较均一,不分化成栅栏组织和海绵组织,所以,禾本科植物叶是等面叶,叶肉内的胞间隙较小,在气孔的内方有较大的胞间隙,即孔下室。
3.叶脉叶脉内的维管束是有限外韧维管束,与茎内的结构基本相似。叶内的维管束一般平行排列,较大的维管束与上、下表皮间存在着厚壁组织。维管束外,往往有一层或二层细胞包围,组成维管束鞘。维管束鞘有两种类型:如玉米、甘蔗、高粱等的维管束鞘,是单层薄壁细胞的,细胞较大,排列整齐,含叶绿体。在显微结构上,这些叶绿体比叶肉细胞所含的为大,没有或仅有少量基粒,但它积累淀粉的能力,却超过叶肉细胞中的叶绿体;水稻、小麦、大麦等的维管束鞘有两层细胞,但水稻的细脉中,一般只有一层维管束鞘。外层细胞是薄壁的,较大,含叶绿体较叶肉细胞中为少;内层是厚壁的,细胞较小,几乎不含叶绿体。禾本科植物叶中的维管束鞘类型不同,一般可作为区分黍亚科(Panicoideae)和早熟禾亚科(Pooideae)的参考依据。
随着科学研究的发展,人们不仅认识到维管束鞘的解剖结构与禾本科植物分类有关,同时,也进一步注意到维管束鞘和它周围叶肉细胞的排列和结构与光合作用的关系。玉米等植物叶片的维管束鞘较发达,内含多数较大叶绿体,外侧紧密毗连着一圈叶肉细胞,组成“花环形”结构(图3-103)。根据近代光合作用途径的研究,这种“花环”解剖结构是碳四(C4)植物的特征。小麦、水稻等植物的叶片中,没有这种“花环”结构出现,并且维管束鞘细胞中的叶绿体也很少,这是碳三(C3)植物在叶片结构上的反映。具有“花环”结构的C4植物叶片中,它的维管束鞘细胞,在进行光合作用时,可以将叶细胞中,由四碳化合物所释放出的二氧化碳,再行固定还原,这样就提高了光合效能。一般认为,C4植物可称为高光效植物,而C3植物为低光效植物。C4和C3植物不仅存在于禾本科植物中,其他一些单子叶植物和双子叶植物中也有发现,如莎草科、苋科、藜科等也有C4植物,而大豆、烟草则属C3植物。
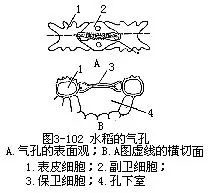

禾本科植物叶脉的上、下方,通常都有成片的厚壁组织把叶肉隔开,而与表皮相接。水稻的中脉,向叶片背面突出,结构比较复杂,它是由多个维管束与一定的薄壁组织组成。维管束大小相间而生,中央部分有大而分隔的气腔,与茎、根的通气组织相通。光合作用所释放的氧,可以由这些通气组织输送到根部,供给根部细胞呼吸的需要。
(三)松针的结构
裸子植物中松属植物是常绿的,叶为针叶,有时称为松针,因而松属植物有针叶植物之称,是造林方面很重要的树种。针叶植物常呈旱生的形态,叶作针形,缩小了蒸腾面积。松叶发生在短枝上,有的是单根的,多数是两根或多根一束的。松叶一束中的针叶数目不同,因而横切面的形状也就不同。例如马尾松(Pinus massoniana)和黄山松(台湾松, P.taiwanensis)的针叶是两根一束,横切面作半圆形,而云南松(P.yunnanensis)是三根一束,华山松(P.armandii)是五根一束,它们的横切面都作三角形(图3-104)。现以马尾松为例,说明针叶内部结构(图3-105,图3-106)。马尾松叶的表皮细胞壁较厚,角质层发达,表皮下更有多层厚壁细胞,称为下皮(hypodermis),气孔内陷,这些都是旱生形态的特征。此外,叶肉细胞的细胞壁,向内凹陷,成无数褶襞,叶绿体沿褶襞而分布,这就使细胞扩大了光合面积。叶肉细胞实际上也就是绿色折叠的薄壁细胞。叶内具若干树脂道,在叶肉内方有明显的内皮层,维管组织两束,居叶的中央。松属的其他种类,有仅具一束维管组织的,因此,按维管组织的束数,而把松属分为两个亚属,即单维管束亚属(subgen.strobus)和双维管束亚属(subgen.pinus)。

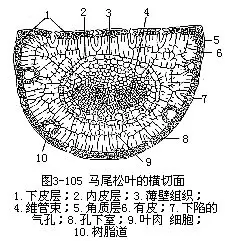

松针叶小,表皮壁厚,叶肉细胞壁向内褶叠,具树脂道,内皮层显著,维管束排列在叶的中心部分等,都是松属针叶的特点,也表明了它是具有能适应低温和干旱的形态结构。
本文标题:第三章 种子植物的营养器官(16)
手机页面:http://m.dljs.net/dlsk/zhiwuxue/59444.html
本文地址:http://www.dljs.net/dlsk/zhiwuxue/59444.html
