(一)顶端分生组织
茎的顶端分生组织和根端的相似。经过顶端分生组织的活动产生了茎的有关结构,包括茎的节和节间、叶、腋芽以及以后转变成生殖(繁殖)结构。茎的顶端分生组织比根的更复杂些,这是因为叶原基的形成和侧枝的发生,都要追溯到顶端分生组织。因此,在讨论茎的顶端分生组织的结构和活动时,也必然要涉及这些侧生器官的起源。
茎的顶端分生组织中夹杂着分化程度不一的组织,这点也和根的相似。在细胞和组织的发育过程中,从分生组织状态过渡到成熟组织状态,是经过由不分化逐渐变为分化的,因而,顶端分生组织的最先端部分,包括原始细胞和它紧接着所形成的衍生细胞,可以看作是未分化或最小分化的部分称原分生组织(promeristem)。在原分生组织下面,随着不同分化程度的细胞出现,逐渐开始分化出未来的表皮、皮层和维管柱的分生组织,也就是它们的前身,可分别称为原表皮(protoderm)、基本分生组织(ground meristem)和原形成层(procambium),总称初生分生组织(图3-34)。初生分生组织的活动和分化的结果,就形成成熟组织,组成初生植物体。总之,茎的顶端分生组织可以说是由原分生组织和初生分生组织组成的。
由于名词应用上存在的混乱,为便于阅读参考书的需要,在这里对生长点、生长锥、茎端、根端、茎尖和根尖等名词分别加以说明。当然,各该名词在不同书中作者所给予的不同解释和范围,也应加以注意掌握,方能有利于正确理解。
茎的顶端分生组织,有时也称生长点(growing point)。这一名词在根中也应用,但严格地讲,生长点不是一个正确的名词,因为生长一般常指细胞分裂,而它确实是分生组织的特征,顶端分生组织也不例外,但细胞分裂却不局限于生长点,有时离生长点较远的部分反而分裂旺盛;生长也指细胞、组织和器官大小的增加,但最显著的增大却不在生长点,而是出现在它的衍生结构部分。生长点有时也称生长锥(growing tip),但生长锥却只用于茎尖上,在栽培学上常应用,一般系指茎尖上最末一个叶原基以上的一段。茎端(stem apex)和根端(root apex)是各自顶端分生组织的同义词,茎端有时也称枝端或苗端(shoot apex)。而茎尖(stemtip)和根尖(root tip)通常指茎或根的顶端分生组织到组织分化接近成熟区之间的一段。如果根冠存在的话;根尖还包括根冠的部分。

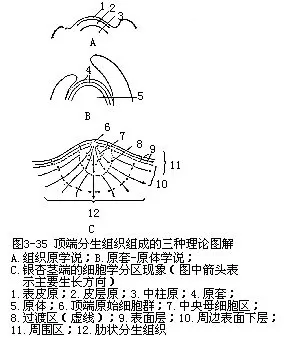
(二)顶端分生组织组成的几种理论
茎的顶端分生组织由许多细胞组成,有着多种方式的排列,在18世纪中叶,就开始引起植物学家的重视,以后陆续提出了不少理论,下面介绍三种理论。
1.组织原学说(histogen theory) 1868年韩士汀(J.von Hanstein)提出了组织原学说。他认为被子植物的茎端是由三个组织区(表皮、皮层、维管柱)的前身,即组织原组成的,每一组织原由一个原始细胞或一群原始细胞发生的。这三个组织原分别称为表皮原(dermato-gen)皮层原(periblem)和中柱原(plerome)(图3-35,A),它们以后的活动能分别形成表皮、皮层和维管柱,包括髓(它如果存在)。由于以后发现茎端不能显著地划分出这三层组织原,所以在茎中是不适用的。但此学说在描述根端组织时比较方便,因而组织原概念,在根中却基本上沿用到现在。组织原学说的提出,使人们对顶端分生组织的认识有了提高,这样,也对顶端分生组织的研究起了积极的推进作用。本章第一节根的顶端组织即按此学说描述。
2.原套-原体学说(tunica-corpustheory)史密特(A.Schmidt)对被子植物的茎端进行研究后,于1924年提出有关茎端原始细胞分层的概念,通常称为原套-原体学说。这个学说认为茎的顶端分生组织原始区域包括原套(tunica)和原体(corpus)两个部分,组成原套的一层或几层细胞只进行垂周分裂(径向分裂),保持表面生长的连续进行;组成原体的多层细胞进行着平周分裂(切向分裂)和各个方向的分裂,连续地增加体积,使茎端加大。这样,原套就成为表面的覆盖层,覆盖着下面组成芯的原体(图3-35,B)。原套和原体都存在着各自的原始细胞。原套的原始细胞位于轴的中央位置上,原体的原始细胞位于原套的原始细胞下面。这些原始细胞都能经过分裂产生新的细胞归入各自的部分。原套和原体都不能无限扩展和无限增大,因为当它们形成新细胞时,较老的细胞就和顶端分生组织下面的茎的成熟区域结合在一起。被子植物中原套的细胞层数各有不同,根据观察,双子叶植物的过半数具有两层,还曾发现有多至四层或五层的,但由于不同学者的划分根据不同,因此,存在着争议。单子叶植物的原套,一般认为只有一层或两层细胞。原套-原体学说认为顶端分生组织(原分生组织部分)的组成上并没有预先决定的组织分区,除表皮始终是由原套的表面细胞层所分化形成的以外,其他较内的各层衍生细胞的发育并不能预先知道它们将形成什么组织,这一点是和组织原学说最大的区别。
3.细胞学分区概念(concept of cytologicalzonation)裸子植物茎端没有稳定地只进行垂周分裂的表面层,也就是没有原套状的结构[除南洋杉属(Araucaria)和麻黄属(Ephedra)外],因此,对于多数裸子植物茎端的描述来讲,原套-原体学说是不适合的。福斯特(A.S.Foster)根据细胞的特征,特别是不同染色的反应, 1935年在银杏(Ginkgo biloba)茎端观察到显著的细胞学分区现象(图3-35,C)。分区的情况是这样的:银杏茎端表面有一群原始细胞即顶端原始细胞群,在它们的下面是中央母细胞区,是由顶端原始细胞群衍生而组成的。中央母细胞区向下有过渡区。中央部位再向下衍生成髓分生组织,以后形成肋状分生组织;原始细胞群和中央母细胞向侧方衍生的细胞形成周围区(或周围分生组织)。中央母细胞区的细胞特征是一般染色较淡,较液泡化和较少分裂。过渡区的细胞在活动高潮时,进行有丝分裂,很像维管形成层。髓分生组织一般只有几层,它的细胞相当液泡化,能横向分裂,衍生的细胞形成纵向行列的肋状分生组织。周围区染色较深,有活跃的有丝分裂,它的局部较强分裂活动的结果,形成叶原基。周围区平周分裂的结果能引起茎的增粗,而垂周分裂则能引起茎的伸长。这种细胞分区现象后来在其他裸子植物和不少被子植物的茎端也观察到,但分区的情况有着较大的变化。对茎端的组织化学的研究,更发现各区细胞不仅形态不同,生物化学方面,如RNA、DNA、总蛋白等的浓度,也有差异,这就反映出分区情况的变化是由于局部区域之间真正生理上的不同。因此,某一植株茎端的分区,在个体发育的不同时期以及不同种之间都可能存在着差异。由于这种分区的研究,以后不再停留在原分生组织的部分,而扩展到衍生区域,因此,茎的顶端分生组织的概念也就扩大。原来将原分生组织和顶端分生组织作为同义词来看,也就不再适合,因而把顶端分生组织的最远端称为原分生组织,似乎更适合些。
关于顶端分生组织组成的理论,还有几种,这里就不一一介绍了。

(三)叶和芽的起源
1.叶的起源叶是由叶原基逐步发育而成的(图3-36)。裸子植物和双子叶植物中,发生叶原基的细胞分裂,一般是在顶端分生组织表面的第二层或第三层出现。平周分裂增生细胞的结果,就促进了叶原基的侧面突起。突起的表面出现垂周分裂,以后这种分裂在较深入的各层中和平周分裂同时进行。单子叶植物叶原基的发生,常由表层中的平周分裂开始。
原套或原体的衍生细胞,都可分裂引起原基的形成。原套较厚时,整个原基即可由原套的衍生细胞发生。否则,叶原基可由原套和原体共同产生。
刚开始发生的侧面突起,是叶原基形成中的开始阶段,通常称为叶原座(leaf buttress),它是整个叶的萌芽,而不是叶的一部分。叶原基出现在顶端分生组织的周围,其相对位置与枝上的叶序相一致。
2.芽的起源顶芽发生在茎端(枝端),包括主枝和侧枝上的顶端分生组织,而腋芽起源于腋芽原基。大多数被子植物的腋芽原基,发生在叶原基的叶腋处。腋芽原基的发生,一般比包在它们外面的叶原基要晚。腋芽的起源很象叶,在叶腋的一些细胞上经过平周分裂和垂周分裂而形成突起,细胞排列与茎端的相似,并且本身也可能开始形成叶原基。不过,在腋芽形成过程中,当它们离开茎端一定距离以前,一般并不形成很多叶原基。
茎上的叶和芽起源于分生组织表面第一层或第二、三层细胞,这种起源的方式称为外起源。不定芽的发生和顶芽、腋芽有别,它的发生与一般顶端分生组织无直接关系,它们可以发生在插条或近伤口的愈伤组织、形成层或维管柱的外围,甚至在表皮上,以及根、茎、下胚轴和叶上。不定芽的起源依照发生的位置,可以分为外生的(靠近表面发生的)和内生的(深入内部组织中发生的)两种。当开始形成时,由细胞分裂组成顶端分生组织,当这种分生组织形成第一叶时,不定芽与产生芽的原结构之间建立起维管组织的连续,而这种连续是由不定芽的分化,和原有的维管组织的相接而形成的。
本文标题:第三章 种子植物的营养器官(10)
手机页面:http://m.dljs.net/dlsk/zhiwuxue/59444.html
本文地址:http://www.dljs.net/dlsk/zhiwuxue/59444.html
