茎的顶端分生组织的活动使茎伸长,这个过程称为初生生长,初生生长中所形成的初生组织组成初生结构。初生生长中,也有增粗,一般是少量的,各种植物间存在着差异。以后茎的侧生分生组织的细胞分裂、生长和分化的活动使茎加粗,这个过程称为次生生长,次生生长所形成的次生组织组成了次生结构。所谓侧生分生组织,包括维管形成层和木栓形成层。多年生的裸子植物和双子叶木本植物,不断地增粗和增高,必然地需要更多的水分和营养,同时,也更需要大的机械支持力,这也就必须相应地增粗即增加次生结构。次生结构的形成和不断发展,就能满足多年生木本植物在生长和发育上的这些要求,这些也正是植物长期生活过程中产生的一种适应性。少数单子叶植物的茎也有次生结构。但性质不同,加粗也是有限的。
(一)双子叶植物茎和裸子植物茎的次生结构
1.双子叶植物茎的次生结构
(1)维管形成层的来源和活动
①维管形成层的来源初生分生组织中的原形成层,在形成成熟组织时,并没有全部分化成维管组织,在维管束的初生木质部和初生韧皮部之间,留下了一层具有潜在分生能力的组织,即维管形成层(以后简称形成层),在初生结构中,它位于维管束的中间部分,即韧皮部和木质部之间,因此,也称为束中形成层(fascicularcambium,图3-51)。
初生结构中,曾提到髓射线,即维管束之间的薄壁组织,在这个组织中,相当于形成层部位的一些细胞恢复分生能力时,称为束间形成层(interfascicular cambium,图3-51)。束间形成层产生以后,就和束中形成层衔接起来,在横切面上看来,形成层就成为完整的一环。从来源的性质上讲,束中形成层和束间形成层尽管完全不同,前者由原形成层转变而成,后者由部分束间薄壁组织细胞恢复分生能力而成,但以后二者不论在分裂活动和分裂产生的细胞性质以及数量上,都是非常协调一致的,共同组成了次生分生组织。
原形成层是由茎(或根)的顶端分生组织前端的原始细胞分裂产生的衍生细胞再分化而成。种子植物的原形成层开始发生时多形成环状一圈,以后随着叶原基的出现,而逐渐分化成束状的维管组织,包括其间的形成层(束中形成层)。由于当前对原形成层转变成束中形成层的过程研究得不多,意见也不一致,在各种植物中转变的过程也相悬殊,有的清楚,有的二者的界限却难以分辨,因之,上述情况只是就一般而言。
原形成层既是束中形成层的前身,为了更好地理解束中形成层的性质,现就原形成层和束中形成层的差异作一扼要的比较:原形成层位于顶端分生组织的下方,细胞较小,细胞质浓厚,各细胞间无大差异,是较均一的组织;束中形成层位于初生木质部和初生韧皮部之间,细胞大小、长短不一,并液泡化,系非均一的组织。原形成层以后本身完全转变成初生维管组织,也不再存在原始细胞;形成层的细胞每次分裂产生两个子细胞,一个分化成维管组织的组成分子或射线;另一个仍保留原来的分生能力并继续分裂和分化,也就是存在着不断更新的原始细胞。
不论束中形成层或束间形成层,它们开始活动时,细胞都是进行切向分裂,增加细胞层数,向外形成次生韧皮部母细胞,以后分化成次生韧皮部,添加在初生韧皮部的内方;向内形成次生木质部母细胞,以后分化成次生木质部,添加在初生木质部的外方。同时,髓射线部分也由于细胞分裂不断地产生新细胞,也就在径向上延长了原有的髓射线。茎的次生结构不断地增加,达一定宽度时,在次生韧皮部和次生木质部内,又能分别地产生新的维管射线(图3-52)。
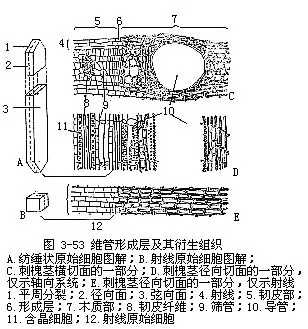
②维管形成层的细胞组成、分裂方式和衍生细胞的发育就形成层的细胞组成来讲,形成层细胞有纺锤状原始细胞和射线原始细胞两种类型(图3-53)。纺锤状原始细胞,形状像纺锤,两端尖锐,长比宽大几倍或很多倍,细胞的切向面比径向面宽,其长轴与茎的长轴相平行。射线原始细胞和纺锤状原始细胞不同,从稍为长形到近乎等径,它们的细胞特征很像一般的薄壁细胞。就纺锤状原始细胞讲,它分裂后,衍生的细胞中有些形成次生韧皮部和次生木质部,但另一些细胞却仍然形成纺锤状原始细胞,始终保持继续分裂的特性,只是这些细胞本身在不断地更新。射线原始细胞也是这样,它的衍生细胞一部分分化形成射线细胞,而另一部分却又继续成为新的射线原始细胞。
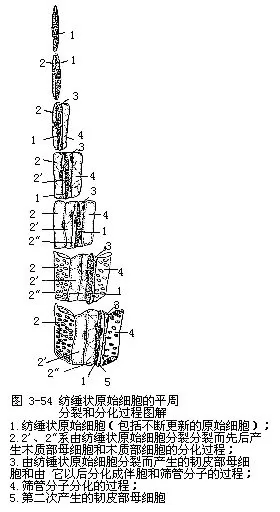
形成层究竟怎样形成次生维管组织和射线呢?关键在于形成层细胞的分裂方式。形成层细胞以平周分裂的方式形成次生维管组织(图3-54)。形成层细胞(即原始细胞,只有一薄层(理论上只是一层细胞),但它活跃地进行分裂时,新的衍生细胞已经产生,老的衍生细胞还在分裂,这时候很难区分原始细胞和它的衍生细胞,特别是衍生细胞在分化成次生韧皮部和次生木质部细胞以前,往往也要进行一次或几次平周分裂,因而通常把原始细胞和尚未分化而正在进行平周分裂的衍生细胞所组成的形成层带(cambial zone),笼统地称为“形成层”,在工作上较为方便。

形成层形成的次生木质部细胞,就数量而言,远比次生韧皮部细胞为多。生长二三年的木本植物的茎,绝大部分是次生木质部。树木生长的年数越多,次生木质部所占的比例越大。十年以上的木质茎中,几乎都是次生木质部,而初生木质部和髓已被挤压得不易识别。次生木质部是木材的来源,因此,次生木质部有时也称为木材。
双子叶植物茎内的次生木质部在组成上和初生木质部基本相似,包括导管、管胞、木薄壁组织和木纤维,但都有不同程度的木质化。这些组成分子都是由形成层的纺锤状原始细胞分裂、生长和分化而成,它们的细胞长轴与纺锤状原始细胞一致,都与茎轴相平行,所以共同组成了和茎轴平行的轴向系统。次生木质部中的导管类型以孔纹导管最为普遍,梯纹和网纹导管为数不多。导管的大小、数目和分布情况,在不同种类植物中,有很大的差异。木薄壁组织贯穿在次生木质部中成束或成层,数量不少,在各种植物的茎中,围绕或沿着导管分子有多种分布方式,是木材鉴别的根据之一。木纤维在双子叶植物的次生木质部,特别是晚材中,比初生木质部中的数量多,成为茎内产生机械支持力的结构,也是木质茎内除导管以外的主要组成分子。次生木质部与初生木质部组成上的不同,在于它还具有木射线。木射线由射线原始细胞向内方产生的细胞发育而成,细胞作径向伸长和排列,构成了与茎轴垂直的径向系统,它是次生木质部特有的结构。木射线细胞为薄壁细胞,但细胞壁常木质化。
生长一年的木质茎内,由于维管形成层活动的结果,已经产生次生维管组织,其中次生木质部已显著,同时在有些植物中,表皮下也出现了木栓形成层,形成了周皮。
形成层向外方分裂的细胞,经过生长和再一二次分裂后,不久就分化成次生韧皮部。次生韧皮部的组成成分,基本上和初生韧皮部中的后生韧皮部相似,包括筛管、伴胞、韧皮薄壁组织和韧皮纤维,有时还具有石细胞。但各组成成分的数量、形状和分布,在各种植物中是不相同的。次生韧皮部中还有韧皮射线(phloem ray)它是射线原始细胞向次生韧皮部衍生的细胞作径向伸长而成,细胞壁不木质化,形状也没有木射线那么规则,这是次生韧皮部特有的结构。筛管、伴胞、韧皮薄壁组织和韧皮纤维由纺锤状原始细胞产生,构成了次生韧皮部中的轴向系统,韧皮射线则构成次生韧皮部的径向系统。韧皮射线通过维管形成层的射线原始细胞,和次生木质部中的木射线相连接,共同构成维管射线(vascular ray)。木本双子叶植物每年由形成层产生新的维管组织,也同时增生新的维管射线,横向贯穿在次生木质部和次生韧皮部内。导管或管胞中的水分,可以借维管射线横向运输到形成层和次生韧皮部;筛管中的有机养料,也可借维管射线横向运输到形成层和次生木质部。维管射线既是横向输导组织,也是储藏组织。从排列方向和生理功能上看,维管射线和髓射线相似,但从起源、位置、数量上看,二者全然不同。维管射线是由射线原始细胞分裂、分化而成,因此,是次生结构,所以也称次生射线(secondary ray),它位于次生木质部和次生韧皮部内,数目不固定,随着新维管组织的形成、茎的增粗也不断地增加。髓射线是由基本分生组织的细胞分裂、分化而成,因此,在次生生长以前是初生结构,所以,也称初生射线(primary ray),它位于初生维管组织(维管束)之间,内连髓部,外通皮层,虽在次生结构中能继续增长,形成部分次生结构,但数目却是固定不变的。
次生韧皮部形成时,初生韧皮部被推向外方,由于初生韧皮部的组成细胞多是薄壁的,易被挤压破裂,所以,茎在不断加粗时,初生韧皮部除纤维外,有时只留下压挤后片断的胞壁残余。
在具双韧维管束的植物中,形成层只存于外韧皮部与木质部之间,以后形成层的活动结果,形成次生结构,而内韧皮部与木质部之间不存在形成层,或存在极微弱的形成层,因而也就不形成次生结构,或形成极少而不显著的次生结构。
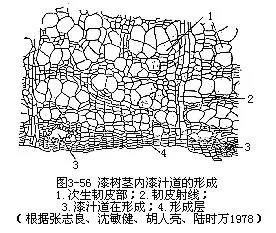
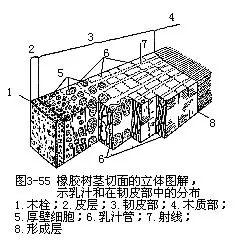
在茎的横切面上,次生韧皮部远不及次生木质部宽厚。这是由于形成层向外方分裂的次数,没有像向内方分裂的次数多,因而,外方新细胞的数量,相应地也就减少。加上次生韧皮部有作用的时期较短,筛管的运输作用不过一二年。当木栓形成层在次生韧皮部发生后,木栓以外的次生韧皮部就被破坏死亡,转变为硬树皮(即落皮层)的一部分,逐年剥落,或积聚在茎干上,因植物种类不同而异。许多植物在次生韧皮部内有汁液管道组织,能产生特殊的汁液,为重要的工业原料(图3-55,图3-56)。例如,橡胶树的乳汁管所产的乳汁,经加工后成为橡胶,漆树的漆汁道所产的漆液,经加工后成各种生漆涂料。不论乳汁管或漆汁道,都分布在次生韧皮部内。此外,有些植物茎的次生韧皮部内,有发达的纤维,可供纺织、制绳、造纸等原料,如黄麻、构树等。
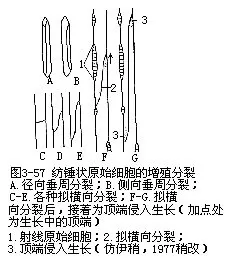
纺锤状原始细胞的分裂,不断地增生次生维管组织,特别是次生木质部,使茎的周径不断增粗。因此,形成层的周径也必然地随着相应扩大和位置外移,才能与次生木质部的不断增长相适应。形成层的周径究竟怎样扩大呢?简单地讲,形成层的原始细胞必须增加,也就是需要有它自身的分裂。形成层增加自身原始细胞的分裂,称为增殖分裂(multiplicative division)。以纺锤状原始细胞的增殖分裂来讲,有以下三种形式(图3-57):径向垂周分裂——一个纺锤状原始细胞垂直地或近乎垂直地分裂成两个子细胞,子细胞的切向生长就使切向面增宽;侧向垂周分裂——纺锤状原始细胞的一侧分裂出一个新细胞,它的生长也同样地使切向面增宽;拟横向分裂(或假横向分裂,pseudo-transverse division)——纺锤状原始细胞斜向地垂周分裂,几乎近似横向分裂,两个子细胞通过斜向滑动,各以尖端相互错位,上面的一个向下伸展,下面的一个向上延伸,产生纵向的侵入生长,也就是正在生长的子细胞插入相邻细胞间,在向前延伸中,各以尖端把另一细胞沿着胞间层处加以分离,这种生长类型称为侵入生长(intrusivegrowth或interpositional growth,图3-57,F-G)。结果两个子细胞成为并列状态,通过生长使形成层原始细胞的长度和切向宽度都能增加。基于上述的三种增殖分裂方式,就可不断地增加形成层的周径,包围整个增大中的次生木质部。
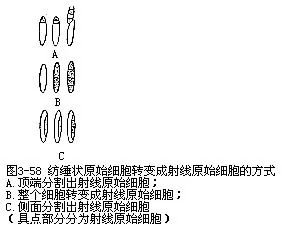
随着茎周径的增粗,相应地次生木质部和次生韧皮部中也不断地分别增生木射线和韧皮射线。这些射线又是怎样产生的呢?由于射线原始细胞分布在纺锤状原始细胞间,因此,射线原始细胞的增殖分裂,也由纺锤状原始细胞的转化来增殖,通常有以下几种形式(图3-58):纺锤状原始细胞的侧向分裂,即在原始细胞中部纵向分割出一部分,形成射线原始细胞;纺锤状原始细胞近顶端横向分割出一个射线原始细胞;纺锤状原始细胞的一半分割成单列射线原始细胞;纺锤状原始细胞的整体分割成单列射线原始细胞;纺锤状原始细胞衰退而逐渐缩短,形成射线原始细胞。一个射线原始细胞可再分裂成一列射线原始细胞;单列射线原始细胞可垂周分裂形成双列,以至形成多列;双列和多列的射线原始细胞也可由于并合而成较宽的。射线原始细胞的增殖分裂和细胞扩大,也对形成层周径的增大起一定的作用。
③维管形成层的季节性活动和年轮
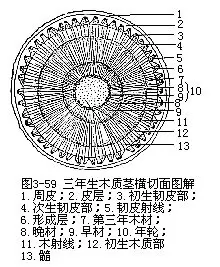
早材和晚材形成层的活动受季节影响很大,特别是在有显著寒、暖季节的温带和亚热带,或有干、温季节的热带,形成层的活动就随着季节的更替而表现出有节奏的变化,有盛有衰,因而产生细胞的数量有多有少,形状有大有小,细胞壁有厚有薄,次生木质部在多年生木本植物茎内,一般比例较大,因此,由于季节的影响,不同时期,它在形态结构上也就出现显著的差异。温带的春季或热带的湿季,由于温度高、水分足,形成层活动旺盛,所形成的次生木质部中的细胞,径大而壁薄;温带的夏末、秋初或热带的旱季,形成层活动逐渐减弱,形成的细胞径小而壁厚,往往管胞数量增多。前者在生长季节早期形成,称为早材(early wood),也称春材。后者在后期形成,称为晚材(late wood,图3-59),也称夏材或秋材。从横切面上观察,早材质地比较疏松,色泽稍淡;晚材质地致密,色泽较深。从早材到晚材,随着季节的更替而逐渐变化,虽可以看到色泽和质地的不同,却不存在截然的界限,但在上年晚材和当年早材间,却可看到非常明显的分界,这是由于二者的细胞在形状、大小、壁的厚薄上,有较大的差异。温带地区因经过干寒的冬季,形成层的活动可暂时休眠,春季湿温,形成层又开始活动,这种气候变化大,形成层的活动差异大,早材和晚材的色泽与质地也就有着显著的区别。
年轮年轮也称为生长轮(growth ring)或生长层(growth layer)。在一个生长季节内,早材和晚材共同组成一轮显著的同心环层,代表着一年中形成的次生木质部。在有显著季节性气候的地区中,不少植物的次生木质部在正常情况下,每年形成一轮,因此,习惯上称为年轮(annual ring,图3-59)。但也有不少植物在一年内的正常生长中,不止形成一个年轮,例如,柑橘属植物的茎,一年中可产生三个年轮,也就是三个年轮才能代表一年的生长,因此,又称为假年轮,即在一个生长季内形成多个年轮。此外,气候的异常,虫害的发生,出现多次寒暖或叶落的交替,造成树木内形成层活动盛衰的起伏,使树木的生长时而受阻,时而复苏,都可能形成假年轮。没有干湿季节变化的热带地区,树木的茎内一般不形成年轮。因此,年轮这一名词,严格地讲,并不完全正确,但已为人们所习用,故本书仍加采用。
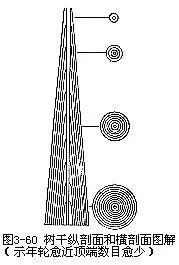
在对于木本植物茎内年轮形成情况了解的基础上,往往可根据树干基部年轮,测定树木的年龄(图3-60)。年轮还可反映出树木历年生长的情况,以及抚育管理措施和气候变化。对年轮反映的树木历年生长情况,结合当地当时气候条件和抚育管理措施的实际,进行比较和分析研究,可以从中总结出树木快速生长的规律,用以指导林业生产。更可以从树木年轮的变化中,了解到一地历年及远期气候变化的情况和规律。有的树龄已达百年、千年之久,以及地下深埋的具有年轮的树木茎段化石,都是研究早期气候、古气候、古植被变迁的可贵依据。
心材和边材形成层每年都不断地产生次生木质部,因而次生木质部也就不断地逐年地大量积累,多年生老茎的次生木质部内外层的性质发生变化,就有心材和边材之分(图3-61)。
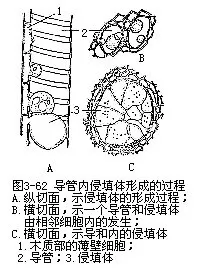
心材(heart wood)是次生木质部的内层,也就是早期的次生木质部,近茎内较深的中心部分,养料和氧进入不易,组织发生衰老死亡,因此,它的导管和管胞往往已失去输导作用,导管和管胞失去作用的另一原因,是由于它们附近的薄壁组织细胞,从纹孔处侵入导管或管胞腔内,膨大和沉积树脂、丹宁、油类等物质,形成部分地或完全地阻塞导管或管胞腔的突起结构,称为侵填体(tylosis,图3-62)。有些植物的心材,由于侵填体的形成,木材坚硬耐磨,并有特殊的色泽,如桃花心木的心材,呈红色,胡桃木呈褐色,乌木呈黑色,使心材具有工艺上的价值。
边材(sap wood)一般较湿,因此也称液材,是心材的外围色泽较淡的次生木质部的部分,也是贴近树皮较新的次生木质部部分,它含有生活细胞,具输导和贮藏作用。因此,边材的存在,直接关系到树木的营养。形成层每年产生的次生木质部,形成新的边材,而内层的边材部分,逐渐因消失输导作用和细胞死亡,转变成心材。因此,心材逐年增加,而边材的厚度却较为稳定。心材和边材的比例,以及心材的颜色和显明程度,各种植物有着较大的差异。
坚实的心材,虽丧失了输导作用,而坚硬的中轴,却增加了高大树木的负载量和支持力。有些木本植物不形成心材或心材不坚,易为真菌侵害,腐烂中空,但边材存在,树木仍然能生活,不过易为暴风雨等外力所摧折。因此,这样中空的高大行道树或观赏树木,就需用加固物质填充已经腐烂中空的部分,以免外力侵袭造成倾倒、坍塌或遭受其他生物的进一步为害。

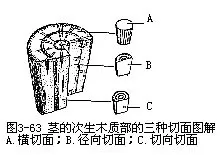
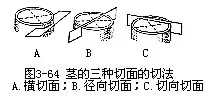
三种切面要充分地理解茎的次生木质部的结构,就必须从横切面、切向切面和径向切面三种切面上进行比较观察(图3-63,图3-64)。这样,才能从立体的形像全面地理解它的结构。横切面是与茎的纵轴垂直所作的切面。在横切面上所见的导管、管胞、木薄壁组织细胞和木纤维等,都是它们的横切面观,可以看出它们细胞直径的大小和横切面的形状;所见的射线作辐射状条形,这是射线的纵切面,显示了它们的长度和宽度。切向切面,也称弦向切面,是垂直于茎的半径所作的纵切面,也就是离开茎的中心所作的任何纵切面。在切向切面上所见的导管、管胞、木薄壁组织细胞和木纤维都是它们的纵切面,可以看到它们的长度、宽度和细胞两端的形状;所见的射线是它的横切面,轮廓呈纺锤状,显示了射线的高度、宽度、细胞的列数和两端细胞的形状。径向切面是通过茎的中心,也就是通过茎的直径所作的纵切面。在径向切面上,所见的导管、管胞、木薄壁组织细胞、木纤维和射线都是纵切面。细胞较整齐,尤其是射线的细胞与纵轴垂直,长方形的细胞排成多行,井然有序,仿佛像一段砖墙,显示了射线的高度和长度。在这三种切面中,射线的形状最为突出,可以作为判别切面类型的指标。
专门研究次生木质部的解剖,也就是研究木材解剖的科学,称为木材解剖学(xylotomy或woodanatomy)。木材解剖学是一门有很大的理论和实践意义的科学,只有对木材的解剖结构有了充分理解,才能很好地判断和比较木材的性质、优劣和用途,从而为林木种类的选择、合理利用,以及为植物的系统发育和亲缘关系等的研究,提供科学依据。
(2)木栓形成层的来源和活动形成层的活动过程中,次生维管组织不断增加,其中特别是次生木质部的增加,使茎的直径不断加粗。一般表皮是不能分裂的,也不能相应地无限增长,所以,不久便为内部生长所产生的压力挤破,失去其保护作用。与此同时,在次生生长的初期,茎内近外方某一部位的细胞,恢复分生能力,形成另一个分生组织,即木栓形成层。木栓形成层也是次生分生组织,由它所形成的结构也属次生结构。木栓形成层分裂、分化所形成的木栓,代替了表皮的保护作用。木栓形成层的结构较维管形成层简单,它只含一种类型的原始细胞,这些原始细胞在横切面上成狭窄的长方形,在切向切面上成较规则的多边形。木栓形成层也和形成层一样,是一种侧生分生组织,它以平周分裂为主,向内外形成木栓和栓内层,组成周皮。

第一次形成的木栓形成层,在各种植物中有不同的起源(图3-65),最通常的,是由紧接表皮的皮层细胞所转变的(如杨、胡桃、榆);有些是由皮层的第二三细胞层转变的(如刺槐、马兜铃);有的是近韧皮部内的薄壁组织细胞转变的(如葡萄、石榴);有些也可由表皮转变而成(如栓皮槠、柳、梨)。木栓形成层的活动期限,因植物种类而不同,但大多数植物的木栓形成层的活动期都是有限的,一般只不过几个月。但有些植物中的第一个木栓形成层的活动期却比较长,有些甚至可保持终生。如梨和苹果可保持6-8年,以后再产生新的木栓形成层;石榴、杨属和梅属的少数种,可保持活动达二三十年;栓皮槠和其他一些种可保持活动达终生,而不再产生新的木栓形成层。当第一个木栓形成层的活动停止后,接着在它的内方又可再产生新的木栓形成层,形成新的周皮。以后不断地推陈出新,依次向内产生新的木栓形成层,这样,发生的位置也就逐渐内移,愈来愈深,在老的树干内往往可深达次生韧皮部。
新周皮的每次形成,它外方的所有的活组织,由于水分和营养供应的终止,而相继全部死亡,结果在茎的外方产生较硬的层次,并逐渐增厚,人们常把这些外层,称为树皮。在林业砍伐或木材加工上,又常把树干上剥下的皮,称为树皮。事实上,前者只含死的部分,后者除死的部分外,却又包括了活的部分,所以,“树皮”这一名词,二者的含义是不同的,往往容易引起混乱。尽管“树皮”并非专业名词,但已为人们所习用,特别是在林业和木材加工方面。因此,对“树皮”这一名词,如能从解剖结构上给予正确的定义,将成为极有用的名词。就植物解剖学而言,维管形成层或木质部外方的全部组织,皆可称为“树皮”(bark)。在较老的木质茎上,树皮可包括死的外树皮(硬树皮或落皮层)和活的内树皮(软树皮)(图3-66,图3-67)。前者包含新的木栓和它外方的死组织;后者包括木栓形成层、栓内层(如果存在)和最内具功能的韧皮部部分。所以在次生状态中的树皮,包括次生韧皮部和可能存留在它外方的初生组织、周皮以及周皮外的一切死组织;有时在初生状态中的所谓树皮,就只包括初生韧皮部、皮层和表皮。

杜仲、合欢、黄檗、厚朴、肉桂等的树皮有着极大的经济价值。过去对一些树皮的采割,常用伐木取皮的方法,这就严重地影响今后的资源。树皮大面积的环剥,长期来被认为由于有机养料运输途径的割断,可以导致整个植株的死亡,所以有“树怕剥皮”之说。但我国农民常对梨、苹果、杏等树适当地进行环剥用以增产,并未发现损害植株。近年来在其他一些树种上进行大面积环剥后,发现仍可正常地再生新树皮的情况。我国植物学工作者*对杜仲(Eucommiaulmoides)进行的剥皮再生的解剖学研究,发现在适当时期剥皮,方法恰当,基本上都能再生出新树皮。他们还发现,剥皮后,近表面的大多数未成熟的木质部分子不久能转变成木栓化细胞,形成保护层。以后在保护层内又逐渐发生木栓形成层,形成周皮。在木质部的内部深层,一些未成熟的木质部分子逐渐转化成新的形成层,由初期不连续的小片,以后变为连续完整的一圈,并不断地分别向内外分化出新木质部和韧皮部。所以,新树皮并不是长期来认为由原来残留的形成层所再生的。这种树皮的再生研究,有着极大的理论上和实践上的意义。
周皮的形成,代替了表皮作为保护组织,但是木栓是不透水、不透气的紧密无隙的组织,那么,周皮内方的活细胞,又怎样才能和外界进行气体交换呢?这里就要介绍一下皮孔,它是分布在周皮上的具有许多胞间隙的新的通气结构,是周皮的组成部分。在树木的枝干表面上,肉眼可见的,具有一定色泽和形状,纵向或横向凸出的斑点,就是皮孔(lenticel)。最早的一些皮孔,往往在气孔下出现,在那部分的深层以后建立了木栓形成层,它和邻近的木栓形成层不同,它的活动不形成木栓,而是产生一些排列疏松、具有发达的胞间隙、近似球形的薄壁组织细胞,它们以后栓化或非栓化,称为补充组织(complementary tissue)。以后由于补充组织的逐步增多,撑破表皮或木栓,形成皮孔。皮孔的形状、色泽、大小,在不同植物上,是多种多样的。因此,落叶树的冬枝上的皮孔,可作为鉴别树种的根据之一。皮孔的色泽一般有褐、黄、赤锈等,形状有圆、椭圆、线形等,大小从1mm左右到2cm以上。就内部结构讲,皮孔有两种主要类型,即具封闭层(closing layer)的和无封闭层的。具封闭层的类型,在结构上有显著的分层现象,这是由于排列紧密、栓化细胞所形成的一至多个细胞厚的封闭层,把内方疏松而非栓化的补充组织细胞包围着。以后,补充组织的增生,破坏了老封闭层,而新封闭层又产生,推陈出新,以此类推,这样,就形成了不少层次的交替排列。尽管封闭层因补充组织的增生而连续遭到破坏,但其中总有一个封闭层是完整的。这种类型常见于梅、山毛榉、桦、刺槐等茎上(图3-68)。无封闭层的类型,在结构上较为简单,无分层现象,但细胞有排列疏松或紧密、栓化或非栓化之分。这种类型常见于接骨木、栎、椴、杨、木兰等的茎上(图3-69)。皮孔也常出现在落皮层裂缝的底部。从皮孔的结构,可以理解它是适应新情况的结构,和表皮上的气孔具有相似的进行气体交换的作用。
2.裸子植物茎的次生结构裸子植物茎和双子叶植物茎比较,裸子植物茎都是木本的,茎的结构基本上和双子叶植物木本茎大致相同,二者都是由表皮、皮层和维管柱三部分组成,长期存在着形成层,产生次生结构,使茎逐年加粗,并有显著的年轮。不同之处是维管组织的组成成分中,有着以下的特点:
(1)多数裸子植物茎的次生木质部主要是由管胞、木薄壁组织和射线所组成,无导管(少数如买麻藤目的裸子植物,木质部具有导管),无典型的木纤维。管胞兼具输送水分和支持的双重作用,和双子叶植物茎中的次生木质部比较,它显出较单纯和原始。在横切面上,结构显得均匀整齐(图3-70)。裸子植物的次生木质部中,也存在着早材、晚材、边材和心材的区分,和双子叶植物茎的次生木质部相同。





(2)裸子植物次生韧皮部的结构也较简单,是由筛胞、韧皮薄壁组织和射线所组成。一般没有伴胞和韧皮纤维,有些松柏类植物茎的次生韧皮部中,也可能产生韧皮纤维和石细胞。
(3)有些裸子植物(特别是松柏类植物中)茎的皮层、维管柱(韧皮部、木质部、髓、甚至髓射线)中,常分布着许多管状的分泌组织,即树脂道。松脂是由松树的树脂道中产生,这在双子叶植物木本茎中是没有的(图3-71)。
(二)单子叶植物茎的次生结构
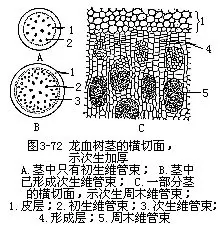
大多数单子叶植物是没有次生生长的,因而也就没有次生结构,它们茎的增粗是由于细胞的长大或初生加厚分生组织平周分裂的结果,在前面的初生结构中已经提过。但少数热带或亚热带的单子叶植物茎,除一般初生结构外,有次生生长和次生结构出现,如龙血树、朱蕉、丝兰、芦荟等的茎中,它们的维管形成层的发生和活动情况,却不同于双子叶植物,一般是在初生维管组织外方产生形成层,形成新的维管组织(次生维管束),因植物不同而有各种排列方式。现以龙血树(Dracaena draco,图3-72)为例,加以说明。
龙血树茎内,在维管束外方的薄壁组织细胞,能转化成形成层,它们进行切向分裂,向外产生少量的薄壁组织细胞,向内产生一圈基本组织,在这一圈组织中,有一部分细胞直径较小,细胞较长,并且成束出现,将来能分化成次生维管束。这些次生维管束也是散列的,比初生的更密,在结构上也不同于初生维管束,因为所含韧皮部的量较少,木质部由管胞组成,并包于韧皮部的外周,形成周木维管束。而初生维管束为外韧维管束,木质部是由导管组成的。

(三)木质茎和草质茎
综上所述,可知茎有木质茎和草质茎之分。裸子植物只有木质茎,双子叶植物既有木质茎,又有草质茎,单子叶植物大多数是草质茎。
1.木质茎在整个植物的进化中,木质茎是较早出现的,裸子植物只有木质茎,就是一个证明。木质茎由于次生结构的发达,木质化的组织占70%以上,质地坚硬而茎干粗大,直径约达50cm的不在少数,最普通的往往也在15cm左右。具木质茎的植物称木本植物,它们的寿命,一般都是几十年到上百年,甚至千年以上。
多数木本植物较老的茎,外表总是粗糙和覆盖着木栓等构成的树皮,树皮上面有通气的皮孔,形式多样。树皮的粗糙和光滑,决定于周皮的结构和发育,以及周皮所分隔成的组织。有木质茎的植物越冬时,芽多数有芽鳞包被着。
幼年的木质茎,当表皮还存在时,表皮内的组织含有叶绿素,能进行光合作用。大多数木本植物,茎外的表皮仅见于初生部分,以后茎加粗,表皮破坏,周皮就代替了表皮,成为次生保护组织,它既不存在含叶绿素的活细胞,光合作用的能力因此也就消失。一般来讲,木质茎上的表皮成熟较早,新枝在第一个生长季内就形成木栓。可是有些木本植物的茎,表皮成熟较迟,茎增粗时,表皮细胞仍能继续分裂,可以延续多年,以适应内部组织的增长,但茎长粗达一定程度,表皮终于被破坏而由周皮所取代,例如樟的茎就是这样。梧桐的表皮生活期较长,能延迟周皮的形成。事实上,大多数一年生左右的木质茎,主要是初生结构时,它的外观和结构都和草质茎相似(参阅图3-38)。随着茎的变老,木质的特征才逐渐发展起来。直径增大,主要是次生木质部产生的结果。
2.草质茎草质茎是由木质茎类型中衍生出来的。草质茎一般柔软、绿色,没有或只有极少量木质化的组织,最多也不超过40%,不能长得很粗,一般停留在初生结构中(图3-73)。大多数单子植物具草质茎。具草质茎的植物称草本植物,寿命往往较短,一般是一年生或二年生,生活期限只有一二个生长季节。有的草本植物有一年生的茎和多年生的根或地下茎,能生活多年,它的茎(地下茎)往往是草质茎,而根是木质的,例如蜀葵、飞燕草、耧斗菜等。
草本植物的茎,如向日葵、棉等,虽产生木质化的组织,但数量少,仍属草质茎。大多数草本植物的茎外部长期存在着表皮,表皮上有气孔。表皮内的组织有叶绿素,因此呈绿色,有进行光合作用的能力。茎的支持作用,依赖厚角组织,厚壁组织和薄壁组织细胞的紧张状态。
草质茎的组织,大部分或完全是初生结构。如和木质茎比较,其维管柱中的维管组织的数量,占较少的比例,这可能是由于维管形成层不发达和活动性下降的缘故。有些双子叶植物,如部分的葫芦科植物,它们的草质茎中,仅有束中形成层,而没有束间形成层。更有些植物如毛茛,不仅没有束间形成层,连束中形成层也不甚发达,活动非常有限,因而次生结构的数量就很少甚至不存在。
必须指出,植物茎的类型不是固定不变的,有些植物生长在某一地区是一年生草质茎,而在另一地区却成为多年生木质茎。例如,番茄和蓖麻在温带较冷的地区是一年生草质茎,而在热带地区却成了多年生木质茎。
本文标题:第三章 种子植物的营养器官(12)
手机页面:http://m.dljs.net/dlsk/zhiwuxue/59444.html
本文地址:http://www.dljs.net/dlsk/zhiwuxue/59444.html
