第一节 从水生到陆生的转变
两栖纲是一类在个体发育中经历幼体水生和成体水陆兼栖生活的变温动物,这个类群绝大多数都是亦水亦陆的种类,也有少数种类终生生活在水中,那是登陆后重新返回水域的次生性现象。现存的两栖动物大多生活在热带、亚热带和温带区域,尤以温暖湿润的热带森林中种类最多,寒带和海岛上的种类却甚稀少,最南分布到新西兰,往北可进入北极圈。迄今为止,南极尚未发现两栖动物的踪进。这些现象表明,温度、湿度和地理障碍等环境因素对两栖动物的发展及其分布范围起着严格的制约作用。
早在距今3.5亿年的古生代泥盆纪,某些具有“肺”的古总鳍鱼曾尝试登陆,并获得初步成功。两栖动物很可能就是在那时由古总鳍鱼类演化而来。最早发现的两栖类化石鱼头螈(Ich-thyostega)与古总鳍鱼类在头骨结构、肢骨等方面均有惊人的相似(图17-1)。但是鱼头螈已经具备了与头骨失去连接的肩带、五趾型的四肢等两栖类的特征。这些古两栖动物大约1.5亿年间,在征服新的陆生环境的同时,迅速地向各方面辐射演化,但以后相继绝灭。现存两栖动物都是从侏罗纪以后才出现的,它们的身体结构及器官机能方面,既保留着原祖的水栖特性,又获得了一系列适应陆地生活的进步特征,居于两者的中间地位。
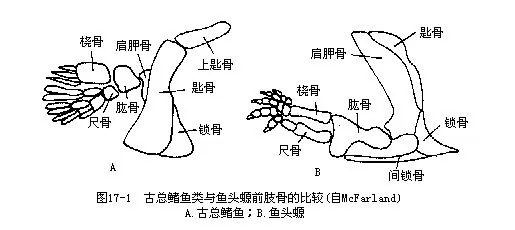
陆地和水域是生存条件具有显著差异的不同环境。水域是由含巨大热能的介质构成,水温的变动幅度不大,一般不超过25℃~30℃,使它能保持在比较稳定的状态。水又是一种密度大于空气千倍的物体,因而尽管它对于动物运动所产生的阻力要比在空气中大得多,但是水具有浮力,能轻而易举地把沉重的动物体承托起来,使它们能在水中遨游。然而,水中的溶氧量只及空气的 1/20,每L水仅含氧3mL~9mL,加以氧的弥散率较低,所以只适于新陈代谢水平低、用鳃进行气体交换和依靠鳍作为运动器官的鱼类生活。
从水生转变到陆生的古两栖动物面临着一系列必须克服的新矛盾:生活介质与气体交换器官的矛盾、浮力消失与动物体承重的矛盾、空气湿度减少与防止体内水分蒸发的矛盾,等等。这些矛盾在古两栖动物的进化过程中,都必须随同环境条件的改变,进行动物体形的相应改造,以及新器官的产生和原有器官的机能转变,否则将导致它们登陆失败而遭受绝灭的命运。在水陆生活转变的许多矛盾中,首当其冲的主要矛盾,就是呼吸器官和陆上运动器官的问题。鱼类在水中生活,由于水能产生浮力,重力对动物的影响较小,借尾、偶鳍和躯体的摆动即可完成运动。两栖动物的成体则不然,它们在空气密度较小的陆地上运动时,不但需要用强健的四肢抵抗重力影响和支撑身体,而且还必须能推动动物体沿着地面移动。正是在这种机能要求的前题下,古两栖动物由酷似古总鳍鱼类的偶鳍发展和形成了适应陆生的五趾型附肢(pentadactyle limb),这是动物演化历史上的一个重要事件。作为鱼类运动器官之一的偶鳍结构比较简单,肩带直接附在头骨后缘,活动的方式和范围受到很大限制,它与鱼鳍之间只有一个单支点,以此作为杠杆,完成单一的转动动作。两栖动物的五趾型附肢与鱼鳍不同,发展了具有多支点的杠杆运动的关节。肩带游离,前肢在摆脱头骨的制约后,不但获得了较大的活动范围,而且也增强了动作的复杂性和灵活性;腰带一方面直接与脊柱牢固地联结,另一方面又与后肢骨相关节,构成支持体重和运动的主要工具,使登陆的目标得以实现。
第二节 两栖纲的主要特征
两栖动物虽已具备登陆的身体结构,但是繁殖和幼体发育仍旧必须在淡水中进行。幼体形态似鱼,用鳃呼吸,有侧线,依靠尾鳍游泳,发育中需经变态(metamorphosis)才能上陆生活,这是两栖纲区别于所有陆栖脊椎动物的基本特征。
一、体形
现存两栖动物的体型大致可分为蚓螈型、鲵螈型和蛙蟾型。蚓螈型的种类外观很像蚯蚓,眼和四肢退化,尾短而不显,以屈曲身体的方式蜿蜒前进,营隐蔽的穴居生活,代表动物有蚓螈和鱼螈等。鲵螈型的种类四肢短小,尾甚发达,终生水栖或繁殖期营水生生活,匍匐爬行时,四肢、身体及尾的动作基本上与鱼的游泳姿势相同,代表动物有各种蝾螈和鲵类。蛙蟾型的体形短宽,四肢强健,无尾,是适于陆栖爬行和跳跃生活的特化分支,也是两栖动物中发展最繁盛和种类最多的类群,代表动物为各种蛙类和蟾蜍。
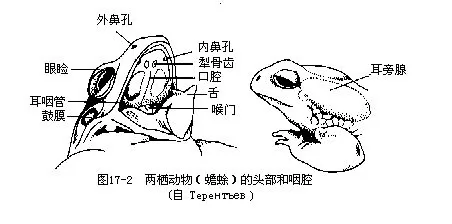
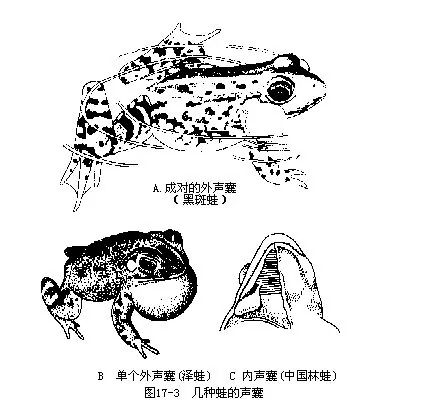
两栖动物的身体分为头、躯干、尾和四肢四部分。头形扁平而略尖,游泳时可减少阻力,便于破水前进。口裂宽阔,颌缘是否有齿视种类不同而异;吻端两侧有外鼻孔一对,具鼻瓣,可随意开闭控制气体吸入和呼出,外鼻孔经鼻腔以内鼻孔开口于口腔前部。大多数陆栖种类的眼大而突出,具活动性眼睑,下眼睑连有半透明的瞬膜(有些鲨鱼已有瞬膜),当蛙、蟾等潜水时,瞬膜会自动上移遮蔽和保护眼球。蛙蟾类的眼后常有一圆形的鼓膜(tympanic membrane),覆盖在中耳(middle ear或称鼓室 tympanic cavity)外壁,内接耳柱骨(columella),能传导声波至内耳产生听觉;中耳还以耳咽管 (eustachian tube)与咽腔连通(图17-2)。雄体的咽部或口角有1~2个内声囊(internal vocal sac,如花背蟾蜍、黑眶蟾蜍、棘胸蛙、中国林蛙、金线蛙等)、或外声囊(externalvocal sac,如黑斑蛙、虎纹蛙、沼蛙、泽蛙、雨蛙、狭口蛙等)。外声囊是由咽壁扩展所成的皮肤囊,充气时可膨胀成泡状或袋状;内声囊位于咽喉腹面或下颌腹面,为皮肤所掩盖,由该处肌肉皱褶向外突出所形成的双壁结构,能发生共鸣作用而扩大喉部发出的叫声。水栖的鲵螈类、大蟾蜍、日本林蛙却无声囊(图17-3),生活于急流山涧内的湍蛙既无声囊,也不发声,而分布在青藏高原的倭蛙和高山蛙不但缺乏声囊,甚至连鼓膜和听骨(耳柱骨)也一并消失,对它们说来,发声和听觉似乎在生活中已经不起任何作用。
颅骨后缘至泄殖孔为躯干部,背面光滑,例如蝾螈、肥螈、雨蛙、姬蛙或粗糙而具瘰粒,例如疣螈、瘰螈、蟾蜍;一些种类常有2条隆起的背褶(dermal plicae),如黑斑蛙、金线蛙、日本林蛙、弹琴蛙;另一些种类却只有长短不一的纵行肤褶(skin fold)或肤嵴(skin ridge),如虎纹蛙、泽蛙等;分布在我国北方的小鲵、山溪鲵、北鲵和爪鲵,体侧均有明显的肋沟(costal groove),尾形侧扁,是鲵螈类的游泳器官。
附肢2对,但鲵螈类中的鳗螈仅有细小的前脚,而蚓螈和鱼螈则四肢已经退化。蛙蟾类的四肢发展很不平衡,前肢短小,4指,指间无蹼(web),主要用作撑起身体前部,便于举首远眺,观察四周;后肢长大而强健,5趾,趾间有蹼,适于游泳和在陆地上跳跃前进。树栖蛙类的指、趾末端膨大成吸盘,能往高处攀爬,吸附在草木的叶和树干上。
二、皮肤
古两栖动物的体表被鳞,有些种类的背部和头部还覆有从真皮鳞演变所成的骨板。现生两栖动物的皮肤裸露并富含腺体,鳞已退化,只有穴居生活的蚓螈类动物在皮下还埋有残存的鳞迹,这是两栖动物区别于其他各纲脊椎动物的主要特征。
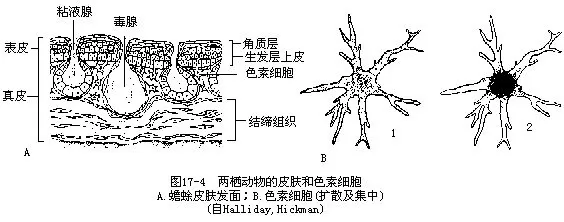
皮肤由表皮和真皮组成,真皮底部有皮下结缔组织,并以此与体肌疏松地相连。表皮是皮肤的外层,含有多层细胞,最内层由柱状细胞构成生发层,能不断地产生新细胞向上推移,由此向外,细胞逐渐变为宽扁形,最外层细胞有不同程度的轻微角质化,称为角质层(stratum corneum),两栖动物皮肤角质化较明显的部位有:蟾蜍头部骨棱的表皮和背上的角质突起或疣粒(wart)、髭蟾上唇边缘的角刺、棘蛙类繁殖前胸部出现的角质刺团、巴鲵掌、跖部的角质鞘和爪鲵指(趾)端的角爪等。角质层细胞从皮肤表面的脱落,就是脊椎动物中常见的蜕皮现象(motting)。表皮中含有丰富的粘液腺,粘液腺为多细胞构成的泡状腺,腺体的分泌部下沉于真皮层,外围肌肉层,有管道通至皮肤表面。粘液腺可借真皮层内的肌纤维收缩,从皮肤开口的腺孔中流出其分泌物,使体表经常保持湿润粘滑和空气、水的可透性,对于减少体内水分散失及利用皮肤进行呼吸都具有重要作用,也是两栖动物通过蒸发冷却用以调节体温的一种途径(图17-4)。雄性狭口蛙和齿突蟾的胸腹部有一大片腺区,其分泌物可使两性在繁殖抱对时牢固地粘贴在一起,不致从背上跌落。位于蟾蜍眼后的耳旁腺(parotid)和皮肤中的毒腺,一般认为是由粘液腺转变而来,能分泌白色、紫红色(盘舌蟾)、褐色(墨西哥蝾螈)或黄色(花背蟾蜍)乳状液的毒浆,内含华蟾毒(cinobu-foxin)、华蟾毒素(cinobufotalin)、华蟾毒精(cinobufagin)等多种有毒成分,对食肉动物的舌和口腔粘膜有强烈的涩味刺激,因而是一种防御的适应。耳旁腺是50~60个椭圆形小毒腺的集合体,每个小毒腺内充满嗜酸性物质,以一短管开口于皮肤表面,腺体的基部有一束平滑肌包绕,控制分泌物的排出。箭毒蛙(Dendrobates)是产于中美洲和南美洲(自哥斯达黎加至巴西)的一类树栖小蛙,因其皮腺内含有剧毒的蛙毒素,故曾被土著居民提取后涂在箭头上,用作自卫和狩猎美洲虎等大型兽类。通常,从一只箭毒蛙分泌物中所提取的蛙毒素,可以用于制作40支毒箭。根据毒性测定,含量为0.00001g的蛙毒素,就足以致一个成人于死地。
真皮厚,居于表皮下方,也分为2层:外层由疏松结缔组织构成,称为疏松层(stratumspon-gosum),疏松层紧贴表皮层,其间分布着大量的粘液腺、神经末梢和血管;内层为致密层(stratumcompactum),内含致密的结缔组织,其中的胶原纤维和弹性纤维呈横形或垂直排列。有些蝾螈在幼体经变态成为能上陆活动的成螈时,真皮内也能出现由多细胞构成的毒腺,藉细管通至体表。
此外,在表皮和真皮中还有成层分布的各种色素细胞(chromatophores),不同色素细胞的互相配置,是构成各种两栖动物体色和色纹的基础。在光线或温度的影响下,色素细胞还能通过其扩展、聚合的形态变化,引起体色改变,由此变成与生活环境浑然一体的保护色,雨蛙和树蛙是两栖动物中具有保护色及能迅速变色的典型代表(图17-4)。
三、骨骼系统
在水栖过渡到陆生的进化中,两栖动物的骨骼发生了巨大变化,获得比鱼类更大的坚韧性、活动性和对身体及四肢的支持作用(图17-5)。
(一)头骨(cranium=skull)
1.宽而扁,脑腔狭小,无眶间隔,属于平底型(platybasic type),枕髁2个,由侧枕骨所形成。
2.骨化程度不高,骨块数目也很少,软骨性硬骨有侧枕骨、眶蝶骨(或单块筛蝶骨)和前耳骨(protic)各一对,而膜性硬骨也只有颅骨背面的鼻骨、额骨、顶骨(或愈合成额顶骨frontoparietal)各一对。颅侧有一块鳞骨(squamosal),颅底由单块副蝶骨(parasphenoid)和一对犁骨构成。鱼类中的许多骨块(如眼眶周围的膜性硬骨)已经消失或仍处于软骨状态。
3.颅骨通过方骨(quadrate)与下颌连接,这种连接方式称自接型(autostylic)。初生颌(腭方软骨和麦氏软骨)趋于退化,由其外包的膜性硬骨(前颌骨、上颌骨和齿骨等)组成的次生颌,代为执行上、下颌的功能。鲵螈类因颧骨(jugal)和方轭骨(quadratojugal)消失,致使颅骨的边缘变得不完整。
4.舌弓背部的舌颌骨移至中耳内,转化成听骨——耳柱骨。
5.幼体时期的鳃弓退化,其残余部分于成体中转变为支持舌和喉部的软骨。
(二)脊柱 在鱼类分化为躯椎和尾椎的基础上,进一步发展成颈椎(cervicalvertebra)、躯干椎、荐椎(sacral vertebra)和尾椎。颈椎一枚,因形状似环又称寰椎(atlas),椎体前有一突起与枕骨大孔的腹面连接,突起的两侧有一对关节窝与颅骨后缘的两个枕髁关节,使头部有了上下运动的可能性;横突不发达,无肋骨;椎骨后端有2个后关节面。荐椎一枚,横突及肋骨大而粗状,外端与腰带的髂骨(ilium)连接,使后肢获得较为稳固的支持。与真正的陆栖脊椎动物相比,因其颈椎和荐椎的数目较少,所以在增加头部运动及支持后肢的功能方面还处于不完善的初步阶段。躯干椎的数目在不同种类之间存在着很大差异(7~200枚)。一般说来,水栖鲵螈类的躯干椎12~16枚,尾椎数大多在20枚以上,原始种类(异鲵Xenobius)的前面几枚尾椎尚留有尾肋的遗迹;半陆生蛙蟾类的进化趋势却是脊柱变短而躯干椎的数目减少至7枚左右,尾椎骨愈合成一根棒状的尾杆骨(urostyle),这是由于水栖生活的躯体波浪状摆动运动方式被半陆栖生活的四肢爬跳运动方式代替的缘故。两栖动物的躯干椎具有陆生脊椎动物椎骨的典型结构,每枚躯干椎均由椎体(除少数种类为双凹型外,大多为前凹型procoelous和后凹型opithocoelous,可增大椎体间的接触面,提高支持体重的效能)、棘突(髓棘)和成对的前关节突、后关节突所组成,因而增强了脊柱的牢固性和灵活性。脊柱两旁,前后相邻2个椎骨之间,有一椎间孔是脊神经向外延伸到躯体各处的必经之孔。
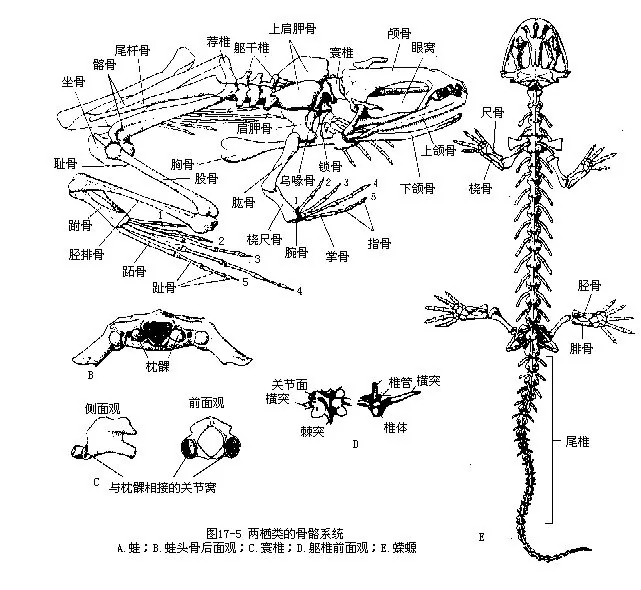
(三)带骨和肢骨 两栖动物的肩带不附着于头骨,腰带借荐椎与脊柱联结,这是四足动物与鱼类的重要区别。肩带脱离了与头骨的连接后,不但可以增进头部的活动性,并且也极大地扩展了前肢的活动范围。蛙蟾类的肩带由肩胛骨(scapula)、乌喙骨(coracoid)、上乌喙骨(epicora-coid)和锁骨(clavicle)等构成,并在胸部正中出现了胸骨(sternum),但与躯干椎的横突或肋骨互不连接。腰带由髂骨、坐骨(ischium)和耻骨(pubis)构成骨盆。鲵螈类的肩带、胸骨和耻骨等大多没有骨化,也缺乏锁骨。组成肩带和腰带的诸骨交汇处,分别形成肩臼(glenoid fossa)和髋臼(acetabulum),与前、后肢相关节。腰带中的耻骨位于髋臼腹面,髂骨和坐骨位于髋臼背面,前者与荐椎两侧的横突关连,这种排列方式,是所有陆生脊椎动物腰带的共性。
两栖动物具有五趾型附肢。一个典型的附肢包括上臂(brachium,股thigh)、前臂(antibra-chium,胫shank)、腕(wrist,跗tarsus)、掌(palm,跖metatarsus)和指(digits,趾digits)等五部分。与之相应的前肢骨为肱骨(humerus)、桡骨(radius)、尺骨(ulna)、腕骨(carpus)、掌骨(metacarpus)和指骨(phalanx)。后肢的五部分分别由股骨(femur)、胫骨(tibia)、腓骨(fibula)、跗骨(tarsus)、跖骨(metatarsals)及趾骨(phalanx)组成。此外,蛙蟾类的拇趾(hullux)内侧还有一个距(calcar)。
四、肌肉系统
两栖动物的肌肉组成了体壁、运动器官和多种内脏器官,并依靠骨骼肌的收缩,产生协调的运动。
两栖动物由水生转变为陆生时,身体和四肢的运动从单一的游泳变得更加复杂,出现了屈背、扩胸、爬行及跳跃等不同形式的活动。因此,与这些运动有关的肌肉都得到了相应的发展。同鱼类相比,两栖动物的肌肉有以下特点(图17-6)。
1.除了幼体(蝌蚪)和鲵螈类外,原始的肌肉按节排列现象在大多数动物的成体已不明显,肌隔消失,大部分肌节由于改变了鱼式运动姿势而发生了愈合或移位,并分化成许多形状和功能各异的肌肉。位于腹部中线两侧的腹直肌(musculus rectus abdominis)和腱划(inscriptio tendine-ae)是显示原有肌肉分节现象的遗迹。然而,在水栖的鲵螈类中,躯干两侧的中轴肌和肌隔均甚发达,肌节与椎骨的数目相当,在扭体摆动运动中,仍保留着它固有的作用。
2.躯干背部的轴上肌由于水平骨隔的位置上移到椎骨横突外侧,因而体积已大为减缩,仅占躯干肌的一小部分。鲵螈类的轴上肌保留着分节状态,蛙蟾类则进一步分化为连接横突之间的横突间肌(musculus intertransversarii)和神经棘之间的棘间肌(musculus interspinalis),并在轴上肌的外侧形成起于头骨基部到尾杆骨前部的背最长肌(musculus longissimus dorsi),其作用是使脊柱向背方弯曲。躯干腹部的轴下肌分化明显,可分为腹直肌和腹斜肌(musculus obliquus ab-dominis)2部分:腹部中线有一条由结缔组织构成的腹白线(linea alba),将腹直肌分隔为左右对称的2份;腹斜肌组成动物体的腹壁,由表及里又分为腹外斜肌、腹内斜肌和腹横肌3层。腹外斜肌呈薄片状,起自背侧的腱膜,肌纤维往后下方走向,覆盖着整个腹腔的后半部,最终在腹直肌的内缘止于白线。腹内斜肌藏于外斜肌下,肌纤维方向指向前上方,正好与腹外斜肌相反。这些肌肉的分化,与其支持腹壁、压缩肺囊和参与呼吸动作有关。腹横肌位于腹壁的最内层,肌纤维呈背腹走向,由耻骨伸往胸骨,有保护腹壁和向前牵拉腰带的作用,以适应两栖类在陆地上的爬行和跳跃运动。
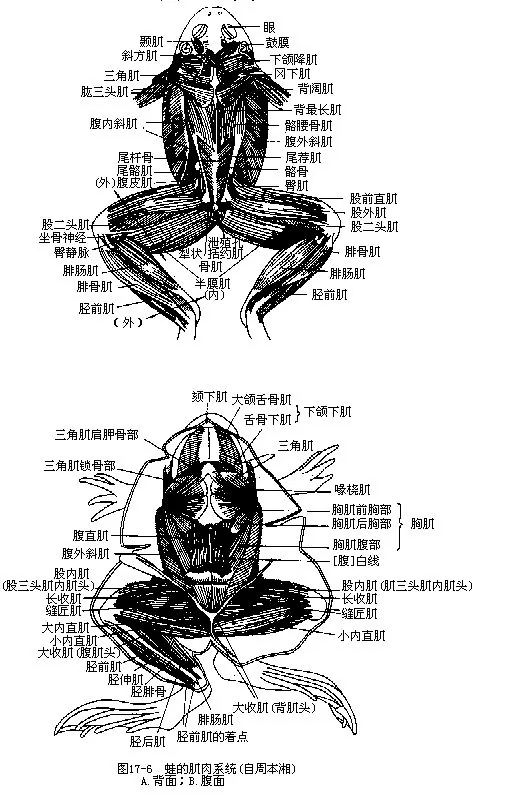
3.在鳍型演变成五趾型附肢的同时,附肢肌也得到了相应的发展,变得强大而复杂,显然这是与它们的肩带不直接连接在头骨上,以及动物上陆后需要具备更有力的附肢支持身体和完成多种形式的运动相联系的。两栖动物具有一些起自头部、躯干、带骨而分布到附肢去的肌肉,称为外来肌,例如,前肢腹侧的胸肌(musculus pectoralis)肌群和背侧的斜方肌(musculus trapezius)、背阔肌(musculus latissimus)、三角肌(musculus deltoideus)等,使前肢与躯干牢固地连接起来而得到巩固。腰带直接与脊柱关连,使后肢得到有力的支持,因此起自躯干、腰带及尾杆骨而止于股部的肌肉数有减少的趋势,主要有梨状肌(musculus piriformis)、臀肌(musculus gluteus)等。
由于四肢分节出现了前肢的肘关节、腕关节和后肢的膝关节、踝关节,因此又分化出许多起点和止点都在附肢骨骼上的肌肉,例如肱三头肌(musculus triceps brachii),腕屈肌(musculusflexor carpi)等前肢肌和胫伸肌(musculus extensor cruris)、缝匠肌(musculus sartorius)、腓肠肌(musculus gastrocnemius)等后肢肌。一般说来,起迄于后肢的肌肉要比前肢更发达,用以加强爬行和跳跃能力。
4.绝大多数两栖动物在幼体时期用鳃呼吸,发育成小动物上陆时鳃已退化消失,鳃弓和鳃肌也转化成支持喉头、舌的软骨,以及节制咽喉部和舌活动的肌肉。
五、消化系统
两栖动物的消化系统包括消化管和消化腺两部分(图17-7)。
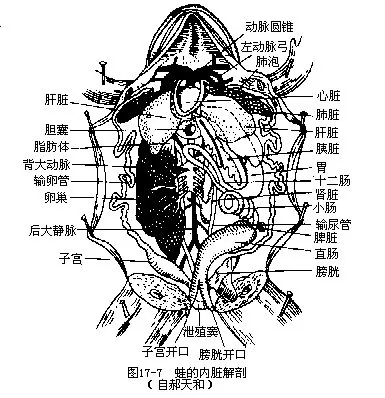
(一)消化管 口腔结构比较复杂,除有牙齿和舌外,还有内鼻孔、耳咽管孔、喉门和食管等开口,分别与外界、中耳、呼吸道、消化管相通。鲵螈类和蚓螈类(鱼螈)的颌缘都有l~2排单尖形的颌齿,而蛙蟾类则无颌齿或仅有上颌齿;齿冠具光泽而坚硬的釉质,齿冠与齿柄以纤维组织相连并形成缢痕,容易断裂,但在功能齿失去齿冠后,齿柄基部的备用齿即可发育长大,代替受损的牙齿,故为多出性的同型齿。此外,口腔顶壁的犁骨上有两簇细小的犁骨齿,这些牙齿并无咀嚼食物的机能,只有咬伤捕食对象和防止食物滑出口外的作用。舌由舌骨(hyoid bone)和舌肌构成,鲵螈类的舌呈垫状,贴于口腔底,活动性较差,舌的后部粘膜内有粘液腺和味蕾;蛙蟾类中除铃蟾等少数种类的舌缘与口腔粘膜相连不能自由伸缩外,其舌根均附着于下颌前部,舌尖游离而有深浅不同的分叉,朝向咽喉部,能迅速翻出口外,粘捕飞行或爬动的昆虫为食。舌的这种特殊结构,是它转变为捕食器官的特化(图17—8)。蛙的口腔中有分泌粘液的颌间腺(intermaxillarygland),位于前颌骨和鼻囊之间,但腺体分泌物无消化功能,只有湿润口腔和食物的作用,并在眼球下沉突向口腔时,协助完成吞咽食物的动作。口腔与咽部似无明显界限。喉门又称声门,位于咽的腹面,是气体出入呼吸道的开孔;咽的背面是食管的开口。

咽部的后方紧缩成管状,即食道的始端,由于颈部分化不明显而食道甚短,其背侧紧贴椎下肌,往后通入胃中。胃位于体腔左侧,前连食道的一端称为贲门(cardia),后连十二指肠的一端称为幽门(pylorus)。胃壁的蠕动,可将食物研碎,胃液能使食物中的蛋白质变成蛋白胨,把脂肪和糖类变成疏松的食物块而有利于小肠中消化液的作用。肠分为小肠和大肠两部分。小肠的主要功能是消化食物和吸收营养,始于幽门,其前段称为十二指肠(duodenum),总输胆管开口于此;后段称为回肠(ileum),几经回旋后,通入宽阔的大肠(intestine crassum)。大肠短而直,故又称直肠(rectum),约为小肠直径的2倍多,两者交界处无括约肌,只有少数蛙蟾类具有活塞状的瓣膜。大肠除吸收水分外,还能聚集不消化的食物,使之后移通入泄殖腔的腹面,再从泄殖孔排出体外。蚓螈类的消化道为一直管,由肠系膜固定在体腔背壁。
各消化器官的内壁,均有发达程度不同和数目不等的纵褶,纵褶下方由粘膜肌层和粘膜下层支持。食管有纵褶12~17条。胃内有纵褶6~14条,其中尤以胃底部的特别粗大;胃壁纵褶基部的粘膜层内,含有大量嗜酸细胞,细胞质中具有嗜红细胞质颗粒,能分泌胃蛋白酶和盐酸,因此被认为是一种未分化类型的“胃腺”。胃壁纵褶中血管丰富,显示两栖动物的胃内可能已初步具有消化吸收的机能。小肠前段的纵褶密集,后段略显稀疏,小肠绒毛为单层柱状细胞;粘膜下层中具有肠腺(intestine gland)。大肠内纵褶甚少,形较低平。(二)消化腺 两栖动物的主要消化腺是肝脏和胰脏。肝脏居体腔的前半部,一般分为2~3叶,胆囊位于肝叶之间或偏右部,是贮存肝细胞分泌胆汁的场所,以数根输胆管与总输胆管相通,并由此将胆汁送入十二指肠远端。胰脏位于胃和十二指肠之间,形状狭长,为不规则的叶状器官(鲵螈类分为背叶和腹叶),有胰管与输胆总管汇合后开口于十二指肠。胰脏分泌胰液与胆汁混合一并注入十二指肠内,在“胃液”初步消化的基础上,进一步消化食物。胰脏不但是一个重要的消化腺,而且还是一个内分泌腺。
六、呼吸系统
两栖动物的幼体和鱼类一样,营鳃呼吸,两者的血液循环方式也几乎完全相同。幼体早期有3对附生于头侧的外鳃 (externalgill),泥螈甚至终生保留着外鳃。外鳃随同幼体发育而被皮肤褶形成的鳃盖所遮掩并逐渐消失,蛙蟾类另以新产生的4对内鳃(internal gill)作为呼吸器官。变态登陆后,内鳃消失,再由咽部腹侧长出一对肺(lung),代替原有鳃的呼吸机能,这是陆栖脊椎动物的重要特征。
呼吸系统包括鼻、口腔、喉气管室(laryngotracheal chamber)和肺脏等。肺是绝大多数种类成体的主要呼吸器官,位于心和肝的背侧,是一对中空半透明和富有弹性的薄壁囊状结构。肺脏内被网状隔膜分隔成许多小室,称为肺泡(alveolus),以此增大肺脏与空气的接触面积;肺泡壁密布着肺动脉和肺静脉的微血管,使动物体的气体交换在肺内得以顺利进行(图17-9)。
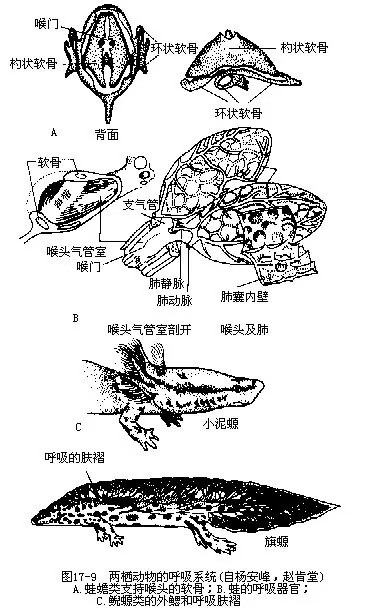
两栖动物肺的结构还比较简单,需要辅助呼吸器官以弥补肺脏摄氧的不足。皮肤薄而湿润,而且在皮下分布着由肺皮动脉(pulmocutaneous artery)分出的皮动脉及肌皮静脉,通过这些皮下血管进行气体交换所得到的氧气,大约相当于肺脏获氧量的2/5。对于鲵类和那些冬季埋身在水底或湿泥中蛰眠的蛙蟾类来说,此时肺呼吸已停止,皮肤便成了代替肺作用的呼吸器官。产于美洲和地中海地区的树螈(Aneides lugubris)是属于无肺螈科(Pleurodontidae)的有尾两栖动物,它们既没有鳃,又缺乏肺脏,完全凭藉分布在皮肤和口腔粘膜下的大量血管进行呼吸。
喉气管室以狭小的裂缝开口于咽部,形成喉门(glottis)。喉气管室由一块环状软骨(cricoidcartilage)和一对构状软骨(arytenoid cartilage)所支持。蛙蟾类在喉门内侧大都附生着一对声带(vocal cord),是2片水平状的弹性纤维带,当空气从肺里冲出时,就会振动声带而发出鸣声。雄性的声带比雌体的发达,并由声囊使鸣声发生共鸣作用,故其鸣声也较雌体更为响亮。鲵螈类一般不能发声。蚓螈类的喉内只有一对杓状软骨,但无环状软骨;具有“C”形软骨环支持的气管,对保证气体在呼吸管中畅通出入具有重要意义;肺呈长囊状,左、右肺不对称,体积差异显著,左肺的长度只及右肺的1/7,但同样都有呼吸作用。
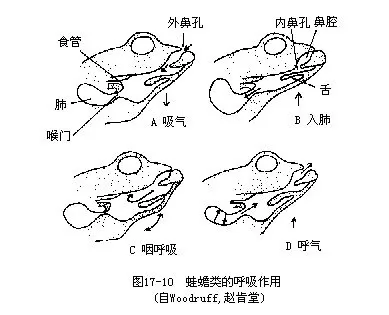
两栖动物的呼吸动作主要依靠口腔底部的颤动升降来完成,并由口腔粘膜进行气体交换,故称口咽式呼吸(bucco-phryngeal respiration)。在一般情况下,外鼻孔的瓣膜张开,喉门紧闭,口底下降而将空气吸入,经内鼻孔到达口内。接着,口底抬升,将空气循原路由鼻孔呼出,此时因喉门始终紧闭而空气不能进入肺脏,只是由口腔粘膜执行气体交换机能。经过口底多次升降颤动后,外鼻孔关闭,口底上举,喉门开启,迫使吸入的空气从口腔进入肺脏,完成气体交换。压迫空气从肺内呼出至口腔的动作,是靠着口底下降,腹壁肌肉收缩和肺脏本身的弹性复原来完成的(图17-10)。
七、循环系统
两栖类由单循环的血液循环方式发展为包括肺循环和体循环的双循环,循环系统包括血管系统和淋巴系统两部分。
(一)血管系统
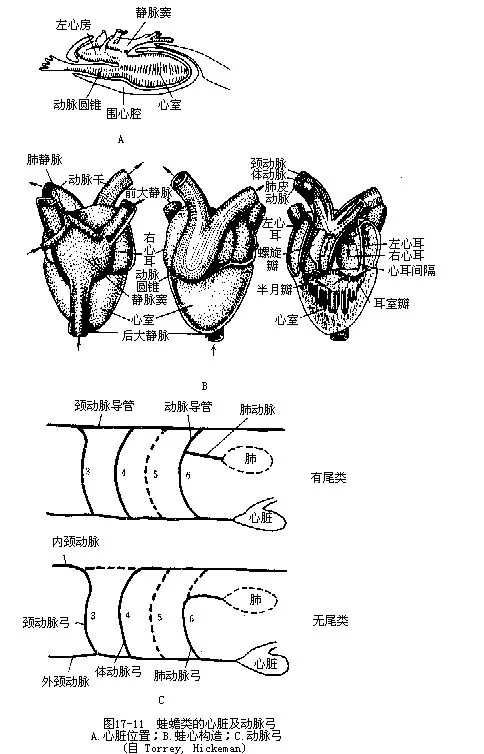
1.心脏 幼体时期的心脏只含有一个心房和一个心室。变态后,心脏的位置由紧挨头部腹面后移至胸腔内,外被围心膜而受到围心腔的很好保护。成体的心脏可分为心室、心房、静脉窦三部分。心室近似三角形,位于心脏后端,壁厚,内有肌质的柱状纵褶,由中央向四周伸展,可在一定程度上缓冲分别由左、右心房流进心室内的多氧血和少氧血的混合。心房位居心室之前,壁薄而色深,内腔被新发生的房间隔分成左、右心房。右心房以窦房孔与静脉窦相通,孔的前、后各有一瓣膜,心房收缩可引起两个瓣膜同时关闭,以防血液发生逆流。左心房的背壁有一孔与肺静脉(pulmonary vien)相通,在肺脏经气体交换后的多氧血,即由此孔进入左心房。左、右两心房分别以房室孔与心室相通,孔的周围有房室瓣(或称三尖瓣),用于阻止血液的倒流。静脉窦是一个三角形的薄壁囊,位于心脏的前端背面,是两条前大静脉(precava)和一条后大静脉(postcava)内的血液流回心脏之前的汇合处。动脉圆锥自心室腹面的右侧发出,与心室连接处有3个半月瓣(valvula semilunaris),瓣的作用也和心脏中的其他瓣膜相同。此外,蛙蟾类的动脉圆锥内还有一纵形的螺旋瓣(spiral valve),能随动脉圆锥的收缩而转动,有助于分配由心脏压出的含氧量不等的血液,循着一定的顺序,分别流入相应的动脉中去。动脉圆锥的前段为腹大动脉,是动脉系统的起点,由此导出3对动脉弓:颈动脉弓(carotid arch)、体动脉弓(systemic arch)和肺皮动脉弓(pulmo-cutaneous arch),它们分别由鱼类中的第三、四、六对动脉弓演变而成,但鲵螈类却保留着第五对动脉弓,并汇合体动脉弓参与背大动脉的组成。蚓螈类只有体动脉弓和肺动脉弓,颈动脉弓由体动脉弓分出至头、脑部(图17-11)。
心脏收缩时,首先由静脉窦开始收缩,将窦内的少氧血注入右心房。接着左、右心房同时收缩,于是右心房内的血液被压入心室中央偏右的一侧,左心房内的血液则被压入心室偏左的一侧。这样,心室中偏右侧为少氧血,偏左侧的为多氧血,中间部分为混合血。心室收缩初期,由于肺皮动脉弓离心室最近,心室右侧的少氧血即率先进入。心室收缩的中期,收缩波从右面移向左边,由于肺皮动脉弓内已充满血液而阻力增高,颈动脉弓基部因有颈动脉腺,阻力也相当大,加以在动脉圆锥收缩时,螺旋瓣往左偏转,关住肺皮动脉的通道,于是心室中部的混合血流入体动脉弓。心室收缩末期,其左侧的多氧血,因受到的压力已达到顶点,便径直注入颈动脉弓,供血给头部及脑。
然而,在X光透视下观察活蛙血液循环的实验却看到:不论是从左心房还是由右心房流到心室的血液,都能流遍心室各部而互相混合,并未见到因心室内存在肌质纵褶使多氧血和少氧血隔开的现象。事实上,许多两栖动物在冬季都停止了肺呼吸,仅靠皮肤呼吸从外界摄取少量氧气,以此维持蛰眠期间的低水平新陈代谢。此时流入左心房的已不是多氧血,而由肌皮静脉流回心室的“多氧血”成了供给全身氧气的主体,这些血液中含氧率极低,几乎全是混合血。由此也可证实,两栖动物在心室内并未严格区分来自左、右心房的多氧血和少氧血。
2.动脉系统 从腹大动脉往前分出一对颈动脉弓,又分为内颈动脉(internal carotid artery)及外颈动脉(external carotid artery),前者运送血液至脑、眼及上颌等处,后者运送血液到下颌和口腔壁。位于内、外颈动脉分叉处的颈动脉腺(carotid gland)是一个压力感受器,用以监测动脉的血压,当动脉血压降低时,颈动脉腺就能发出兴奋传导到延脑的心血管调整系统。
左、右体动脉弓弯向背侧,在分出锁骨下动脉(subclavian artery)至前肢及食道后,便汇合成一条背大动脉,往后延伸并发出动脉分支到内脏各器官及后肢。蚓螈类因无四肢而缺乏锁骨下动脉及髂动脉。
左、右肺皮动脉弓各分为2支,一支是肺动脉(pulmonary artery),通至肺脏,在肺壁上分散成毛细血管网,另一支为皮动脉(cutaneous artery),行至背部皮下,也分散成毛细血管网。肺和皮肤都是两栖动物进行气体交换的器官。
3.静脉系统 心脏以静脉窦接受前大静脉和后大静脉的血液,通过右心房流入心室。肺静脉经左心房进入心室。前大静脉代替了鱼类的前主静脉,汇集由头部和躯干前段的外颈静脉(external jugular vein)、内颈静脉(internal jugular vein)和来自前肢的血液、肌皮静脉(musculo-cutaneous vein)等。后大静脉代替了鱼类的后主静脉,收集来自躯干后段和消化系统返回心脏的血液。尾和后肢的静脉在前行中分为两对,一对沿肾脏的外缘成肾门静脉(renal portal vein),进入肾脏,分成许多细小血管,再次汇集成数条肾静脉(renal vein),由两肾之间通出,与来自生殖腺的生殖腺静脉(genital vein)一起,将血液送入后大静脉;另一对是盆骨静脉(Pelvic vein),在腹壁中央合并成一条腹静脉(abdominal vein),其血液往前注入肝门静脉。从胃、肠、脾、胰等器官所来的静脉合成肝门静脉,进入肝脏,再由肝脏发出一对肝静脉通入后大静脉,最后将血液汇入静脉窦(图17-12)。
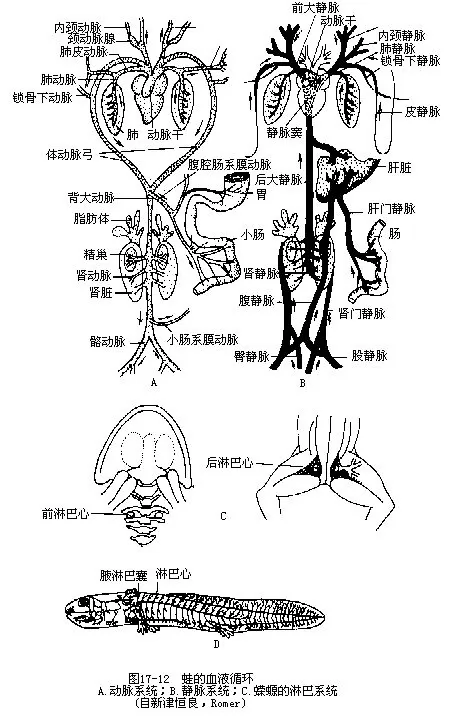
(二)淋巴系统 从两栖类开始出现比较完整的淋巴系统,几乎遍布皮下组织,这与防止皮肤干燥和进行皮肤呼吸有关。包括淋巴管、淋巴腔(lymphatics saccus)和淋巴心等结构。
两栖动物自血管渗出的组织液和从组织细胞内渗出淋巴液的活动较其它脊椎动物更为旺盛而且血压较低,有发达的淋巴心以助淋巴液回心,通常蛙蟾类有淋巴心2对,鲵螈类约16对,蚓螈类甚至可多达百余对。淋巴腔由淋巴管膨大所成,尤以蛙类皮下淋巴腔最为发达。
两栖动物由于不完全的双循环,动脉血液中的含氧量不充分,造成组织细胞中物质的氧化效率不高,新陈代射甚为缓慢,产生的热量少,不足以抵消所丧失的热量,加上没有良好的保温条件,也不具备完善的体温调节机制,因而不能维持恒定的体温,在很大程度上随环境温度而变化,主要借吸收太阳热能来提高体温,所以称为变温动物(poikilothermal)或冷血动物,也叫外温动物(ectothermal)。冬季严寒时,需寻觅适宜的地点入蛰冬眠,此时不吃不动,新陈代谢降到最低水平,依靠夏秋活动期大量摄食积累在体内的脂肪体供给能量。翌年春末夏初出蛰活动。热带地方的两栖动物在酷热的夏季,由于气温太高、环境干旱和食物匮缺,常钻到土层深处湿度较大的洞穴里进行夏眠,暂避不宜的生活条件。两栖动物于夏眠期间,淋巴液和血液中的盐离子浓度有所增加,以此提高体内的渗透压而减少失水。
八、排泄系统
两栖动物的排泄器官是肾脏、皮肤和肺等,但以肾脏最重要,大量的尿液都是在肾内滤出。鲵螈类的肾脏是一对长扁形的带状器官,蛙蟾类是一对结实的椭圆形分叶器官。肾小球的滤过机能强,每天从血液中滤出的水分可达动物自身体重的1/3,因而对于水栖种类维持动物体的内环境恒定,具有十分重要的意义。
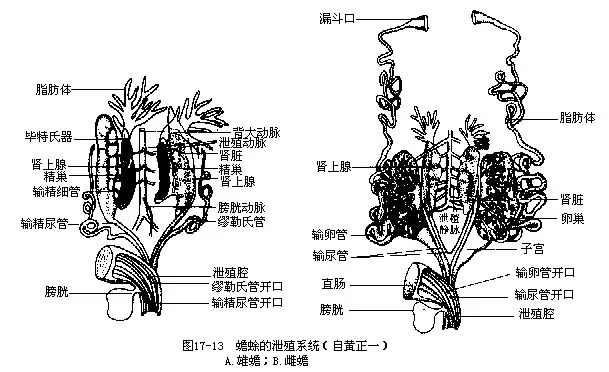
左、右肾的外缘各连接一条输尿管,即中肾管,分别通入泄殖腔的背面。雄体肾脏的前部缩小并失去泌尿功能,由一些肾小管与精巢伸出的精细管相连通,并借道输尿管运送精子,因此兼有输尿管和输精管的用途。雌体的肾脏及输尿管只有泌尿输尿作用,与生殖系统无任何联系。蛙蟾类有一体积较大而薄壁的膀胱,由泄殖腔的腹壁突出所形成,故称泄殖腔膀胱(cloacal blad-der),是暂时贮存尿液的器官。膀胱与输尿管并不直接相通,肾脏滤泌产生的尿液经输尿管先导入泄殖腔并倒流到膀胱里。当膀胱充满尿液后,由于膀胱受压收缩,以及伴随着泄殖孔的张开,才将尿液排出体外(图17-13)。
肾脏除了泌尿功能外,还有调节体内水分和维持渗透压的作用。对于鲵螈类和蛙蟾类来说,淡水是一种含盐值为0.2%~0.5‰的低渗液,外界的大量水分可不断地通过皮肤渗入体内,因而必须依赖过滤机能很强的肾小球及时滤泌成尿,排除潴留在血液中的过量含水,以维持动物体内的水分平衡。如果两栖动物及其幼体和胚胎接触到含盐值超过16‰的水源或土壤时,过多的盐分便可经皮肤进入体内,改变体液的正常浓度,从而造成动物和幼体死亡。
两栖类的膀胱重吸收水分的机能使体内水份的保持得到了加强,但是这种节水作用仍不足以抵偿由于体表蒸发所造成的大量失水。这就决定了两栖动物虽能上陆生活,却不能长时间地远离水源,也是干旱的荒漠地带缺乏两栖动物和温湿多水地区种类繁多的原因之一。陆地上冬眠的种类,蛰眠期间完全依靠皮肤的渗透,从土壤中吸收水分,获取维持生命活动的最低需水量。
九、神经系统
两栖动物神经系统的发展水平与鱼类比较接近,但是随同上陆生活的环境变化及其所受到的影响,使它们的神经系统有了某些进步性的变化。
(一)脑 分五部分,分化程度不高,排列在同一个平面上,并不形成明显的脑曲。大脑的体积增大,往前延伸成2个小形的嗅叶;大脑半球之间以矢状裂相隔,位于脑内的左、右侧脑室,称为第一、第二脑室,2个脑室藉室间孔相通,侧脑室向前一直伸展到嗅叶中,往后与间脑内的第三脑室相通。大脑半球不仅在腹部和侧面保留着神经细胞构成的古脑皮(paleopallium,或称旧脑皮),并且在顶部也发生了零星的神经细胞,称作原脑皮(archipallium),然其机能仍与嗅觉有关。
间脑顶部呈薄膜状,上盖富有血管的前脉络丛,由背面正中伸出一个不发达的松果体,松果体的内腔与间脑中的第三脑室相通,钝口螈(Ambystoma mexicanum)的松果体能产生糖原,输送到脑室的脑脊液内。间脑背侧部的壁厚,称为视丘或丘脑(thalamus),视丘的前下方为下丘脑(hypothalamus),包括视交叉、脑漏斗及脑垂体等。
中脑的背部发育成一对圆形的视叶,腹面增厚为大脑脚(crus cerebri),既是两栖动物的视觉中心,也是神经系统的最高中枢。左、右视叶内皆有宽大的中脑室,两室彼此相通,并以中脑导水管分别与第三、第四脑室沟通。
小脑略呈狭带状,横跨于延脑菱形窝的前缘,紧贴在视叶之后,这种状况与两栖动物的运动方式简单有关。延脑内有一个三角形的第四脑室,与脊髓的中央管相通,脑室顶壁下陷,形成菱形窝,盖有后脉络膜(图17-14)。
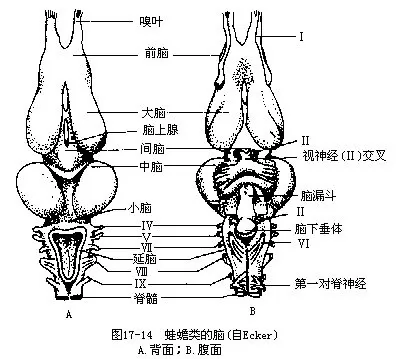
(二)脊髓 除有背正中沟外,还具有脊椎动物中首次出现的腹正中裂(fissura mediana vent-ralis)。由于四肢发达及运动机能的增强,附肢肌完全消失了分节性,并促使脊髓在肱部和腰部发展成2个膨大部分,即颈膨大和腰膨大。
(三)脑神经 10对,但古两栖类的头骨上却有一个舌下神经孔,有人推测可能是该神经退化后的遗迹。
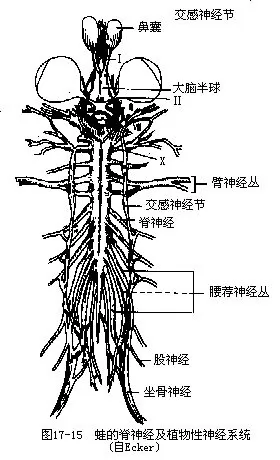
(四)脊神经 其对数因动物类别不同而有很大差异,其中第一对脊神经由寰椎和第二椎骨之间的椎间孔穿出,其分支往前分布到舌肌及部分肩肌上,支配舌的运动。有些脊神经集合成臂神经丛和腰荐神经丛,分别进入前、后肢。
(五)植物性神经系统 较鱼类更为进化,但仍以交感神经为主。交感神经的主体是一对纵行于脊柱两侧的交感神经干,由神经将一系列交感神经节串连而成。交感神经节以交通支与脊神经相连,同时还发出交感神经分布到内脏各器官(图17-15)。副交感神经出现于中枢神经的前段和后段,前段的中枢位于中脑和延脑,副交感神经纤维伴随着第三、七、九、十对脑神经同行,分布到眼、口腔腺、血管和内脏各器官;后段的中枢位于脊髓的荐部,由此发出数对副交感神经,分布到盆腔内的脏器。这些内脏器官同时接受交感神经及副交感神经的支配并以其互相拮抗的作用维持正常生理机能。
十、感觉器官
(一)侧线器官 两栖动物的幼体都具有侧线,由许多感觉细胞形成的神经丘所组成,用作感知水压的变化,幼体变态后侧线消失殆尽。但是分布在欧洲的洞螈、美洲的泥螈、鳗螈、两栖螈及亚洲的大鲵、肥螈、东方蝾螈等水栖鲵螈类,头躯部始终保留着侧线器官和侧线神经,其构造与鱼类极为相似(图17-16)。

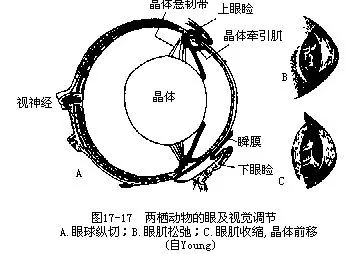
(二)视觉器官 视觉器官已初步具有与陆栖相适应的特点。半陆生的蛙蟾类的眼高踞头的背侧,也可深陷至眼眶内,有能活动的眼睑和瞬膜,还有泪腺(lachrymal gland)和哈氏腺(Harderion gland),这些结构及腺体分泌物都能使眼球润滑,免遭伤害和干燥,有利于陆地生活。大鲵和泥螈的眼小,因其角膜与皮肤愈合而眼睑不能活动,也无泪腺。大多数种类的眼球具有凸出的角膜,晶体近似圆形而稍扁平,以悬韧带固着于眶壁上。由于晶体和角膜的间距较鱼类远,因而适于观看较远处的物体。晶体的腹面(鲵螈类)或背腹面(蛙蟾类)有一块小形的晶体牵引肌,收缩时能将晶体拉向前移和改变其弧度,调整视觉的成像焦距,使之由远视转变成适于近视。此外,在脉络膜和晶体之间还有一些辐射状排列的脉络膜张肌,可能相当于陆生脊椎动物的睫状肌(ciliary muscle),有协助晶体牵引肌调节视觉的作用(图17-17)。总的说来,两栖动物的视觉调节能力不强,视觉调节方式也不同于改变晶体形状的陆生脊椎动物,所以它们在陆地上还只能说是近视动物,只有当潜入水中时,角膜由凸变平,才可能适当地增阔视野。
(三)嗅觉器官 两栖动物的鼻腔内壁衬有褶襞状的嗅粘膜(olfactory mucous membrane),分布在嗅粘膜上的嗅神经往后通至嗅叶,司嗅觉,因此鼻腔开始了兼有嗅觉和呼吸的两重机能。
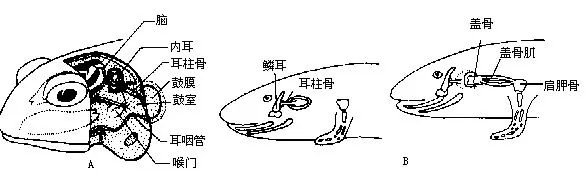
(四)听觉器官 两栖动物在由水生到半陆生的转变过程中,发生了极其深刻的变化。内耳球状囊的后壁已开始分化出雏型的瓶状囊(听壶lagena),有感受音波的作用。因此,两栖动物的内耳除有平衡感觉外,还首次出现了听觉机能。适应在陆地上感受声波而产了中耳(middleear),中耳腔又名鼓室(tympanic cavity),由胚胎的第一对咽囊演变而来。中耳腔内有一枚与鱼类舌颌骨同源的耳柱骨,两端分别紧贴内耳外壁的椭圆窗和鼓膜内面的中央,将鼓膜所感受的声波传入内耳,通过听神经传导到达脑,产生听觉。口腔以一对耳咽管(eustachin tube)与中耳腔相通并进入空气,使鼓膜内、外的受压趋于平衡,防止鼓膜因受剧烈的声波冲击而造成震裂。
鲵螈类和蚓螈类无中耳腔,但有发达的耳柱骨,其外端与鳞骨相关节,通过颌骨可将声波的振动传送到内耳。此外,绝大多数种类的椭圆窗外还有一块平板状的盖骨(operculum),以一小形的盖骨肌(operculum muscle)连接肩带,盖骨肌拉紧时,地面的振动可由附肢经这块肌肉传导到盖骨而达到内耳(图17-18)。
十一、生殖系统
(一)生殖器官 雄性有一对卵圆形(蛙)、长柱形(蟾)或分叶状(蝾螈)的精巢。精液通过输精小管(vassa efferentia)与肾脏前部的肾小管连通,然后借道输尿管进入泄殖腔而排出体外,故名输精尿管。蛙蟾类在繁殖期间,输精尿管的末端膨大成贮精囊(seminal vesicle),用作贮存精液,过了生殖季节则缩小恢复正常。此外,它们还保留着细小而明显的输卵管,这是退化状态的缪勒氏管。除蚓螈类、蝾螈科和尾蟾(Ascaphus truei)等少数种类外,均营体外受精。蚓螈类的泄殖腔长,能突出体外成插入器,将精液直接输入雌体的泄殖腔内,可看作是一种交配器。生活在美国西北部流水中的尾蟾因雄体具有一尾状突而得名,泄殖孔位于尾状突的末端腹面,繁殖季节,雌雄相向抱对,尾状突插入雌蟾泄殖腔孔内,以保证精子与卵相遇。非洲东部的泳蟾(Necto-phrynoides vivipara)虽无交配器,但也属于体内受精的动物,受精卵可在雌蟾的子宫内发育成蝌蚪后产出。分布于欧洲的黑螈(Salamandra atra)和火螈(Salamandra salamandra),受精卵在输卵管内发育,直到形成蝌蚪或小蝾螈时才产出。
雌体有一对囊状结构的卵巢,囊内常含有许多圆形的卵,卵巢和卵的大小、颜色随季节及发育状况而不同。低温蛰眠是大蟾蜍等卵子成熟的决定因素,卵成熟后由卵巢进入腹腔,通过腹腔膜上的纤毛活动和腹肌收缩而进入输卵管前端的漏斗。卵在输卵管内向远端移动的行程中,包上由管壁分泌的胶质,形成卵胶膜,最后到达输卵管扩大的子宫部,于两性配对或抱对后与雄性的精液同时排至水中,完成体外受精。左、右输卵管后部合并为一(蟾)或各自开口于泄殖腔的背壁。动物学家的实验证明了输卵管分泌物是两栖类成熟卵子受精必不可少的物质条件。
蛙蟾类生殖腺的前方都有一对黄色的指状脂肪体(fat bodies),是供给生殖腺发育所需的营养结构。通常,蛰眠期前的蛙和蟾蜍由于摄食旺盛,体内都积贮了丰富的营养物质,因而脂肪体也显得十分粗大。当进入生殖细胞迅速生长发育的繁殖季节,脂肪体被大量消耗而萎缩得很小。此外,蟾蜍属和南美洲的短头蟾科种类的生殖腺前缘都附生着一个形状各异的毕特氏器(Bid-der’s organ),由蝌蚪生殖腺前部膨大部分经变态后形成。毕特氏器相当于退化卵巢的残余部分,内含无数尚未分化或发育不完全的卵细胞。摘去蟾蜍的精巢,可导致毕特氏器发育成具有产卵功能的卵巢,有人认为这种改变并非性逆转现象,而是由于去势后退化卵巢消除了雄性激素的抑制作用所产生的结果。
两栖动物的两性差异一般表现在雄性蛙蟾类的身体略小于雌性,而色斑有区别的种类在我国以花背蟾蜍最明显。吻端的形状在大树蛙的两性间也有显著差异。雄性鲵螈类的肛部隆起呈椭圆形,肛裂较长,内壁有乳白色小疣(大鲵)以及有浅灰色(东方蝾螈)或黑色的绒毛状乳突(肥螈)。繁殖季节内,因受性激素的影响,不少种类的雄性会出现各种形式的副性征:峨嵋髭蟾和棘腹蛙等蛙的前臂因加强抱雌能力变得粗壮而有力,几乎超过正常状态的好几倍。前肢内侧第一、第二指的基部局部隆起成婚垫(nuptial pad),有些种类如铃蟾、花棘蛙(Rana maculosa)、西藏蟾蜍(Bufo tibetanus)的第三指上也有婚垫,垫上富有粘液腺或角质刺、用于加固抱对作用。此外,髭蟾上颌背缘的黑色角刺、棘蛙类胸腹部的腺体及疣刺、棘肛蛙泄殖腔内壁的细棘、隆肛蛙肛部皮肤呈方形的囊状突起,以及旗螈(Triturutus cristatus)背面出现的帆形肤褶等,也都是两栖动物特有的雄性标志(图17-19)。


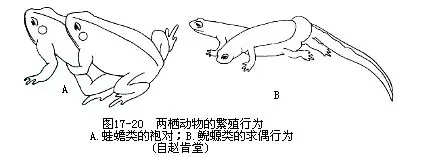
(二)生殖方式 绝大多数种类的繁殖期都在春夏之际,而中国林蛙却于冰雪初融的3月开始抱对,崇安髭蟾和绿臭蛙(Rana margaratae)则可迟至深秋11月或冬季进行产卵。抱对现象是蛙蟾类在产卵前必不可缺的繁殖行为,当雄性一旦追逐到雌体后,鸣声便嘎然而止,用前肢紧紧抱住异性的腋下而蹲伏于其背上。抱对可持续6~8小时,甚至长达一天或数日之久,其生物学意义显然是与刺激两性同步排精产卵和提高受精率具有密切关系。鲵螈类在繁殖期无抱对行为,也不鸣叫招唤异性,但雄体常表现出某些求偶动作:东北小鲵的两性彼此靠拢,结伴游泳,雄体屡屡下沉到雌鲵腹下来回窜游,或以尾部卷绕对方;雄性东方蝾螈常围绕雌体游动,以吻碰触其泄殖孔,尾部不停地弯曲抖动,这种动作可反复多次,甚至持续几小时(图17-20)。
除泳蟾、尾蟾和几种蝾螈营卵胎生外,均以产卵方式进行繁殖。通常雌体都是一次性将所怀的成熟卵全部产出。东方蝾螈,它们在整个繁殖期内每天产一枚受精卵,而泽蛙每年可产卵4~5次,每次的产卵量也比较少。卵及包裹在卵外的胶质膜或胶质囊,其大小、颜色和形状都因动物种类而异:大鲵的卵带成念珠状,小鲵呈圆筒状,蟾蜍的胶质膜连成长条状,青蛙和雨蛙的卵聚成团块状,锄足蟾则集成片状,但蝾螈和铃蟾则产单生卵。两栖动物的卵为多黄卵,动物极含有丰富的细胞质,表层内有黑色素微粒而呈现深褐色或黑色,有利于吸收日光,植物极主要包含卵黄,但无色素,故成灰白或浅黄色。生活在平原地区的蛙蟾类,卵径为1.5mm~3.0mm,而姬蛙的卵形甚小,直径仅0.8mm~l.0mm,栖息于山地溪流中的高山型种类(角蟾、髭蟾、齿突蟾、棘蛙、臭蛙、湍蛙等),卵径增大,直径为3.5mm~4.0mm,最大卵径可达5mm(棘胸蛙),卵乳白色(臭蛙)或灰色(棘蛙等),似与其产卵在石块下不易受到阳光照射有关,树蛙和弹琴蛙能在树叶或土坑内产卵,卵呈现象牙色、浅黄色、浅绿色或灰绿色。
(三)胚胎发育和变态卵外有胶质膜2~3层,豹蛙甚至可达5层之多,胶质膜轻而薄,可被精子头部含有蛋白酸酶所分解和穿透,与卵子结合成受精卵,并吸水膨胀漂浮水面,以利充分接受光照和积贮发育所需的热量。此外,胶质膜还有促进精子正常受精、保护受精卵和使胚胎免遭污染、机械损伤、低渗影响、病原体侵入及水生动物吞食等作用(图17-21)。

![]()
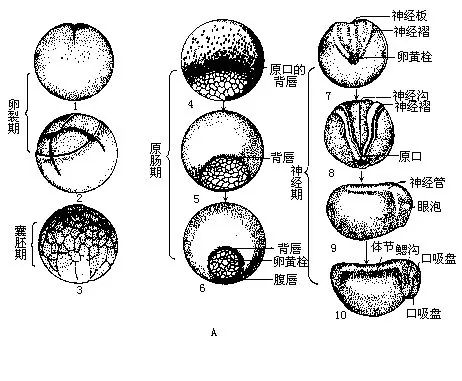
卵在受精后2~4小时便开始分裂。受精卵因内含的卵黄分布不均匀而进行不完全卵裂,动物极和植物极的细胞分裂是非等速进行的。这种卵裂形式与文昌鱼的完全卵裂及鱼类的盘裂不同。受精卵在细胞分裂的增殖初期,称为卵裂期,动物极的细胞以比植物极细胞更快的速度进行有丝分裂,并形成最早的胚胎——囊胚(blastula)。囊胚内具有一个充满液体的囊胚腔。
不间断的细胞分裂,使较小而数量众多的动物极细胞开始向下外包到植物极细胞的表面。同时,植物极细胞也相应地移动和内陷,最后围成原肠腔(archenteron),囊胚腔因受挤消失而被原肠腔所取代。这时胚胎发育进入原肠期,胚胎称为原肠胚(gastrula)。那些内陷的植物极细胞叫作内胚层细胞,外包在胚体表面的动物极细胞称为外胚层细胞。
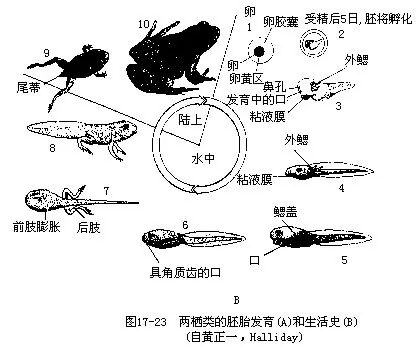
发生外包的动物极细胞后来也由原口处向胚内卷入,即中胚层细胞,其中一部分移到原肠背面形成脊索中胚层,棒状结构的脊索即由此发展而来;其他细胞则转移到胚内的背侧和腹面,以后逐步构成体节、脊柱和体腔等。原肠期开始出现三胚层,不同胚层的细胞已分别聚集在特定位置,为动物体的各种组织、器官和系统的构成作好了准备。原肠期结束时,原口缩小成裂缝状,同时胚胎背面的外胚层细胞又形成神经管,接着下沉至胚内,并为皮肤所覆盖,而其它器官也随之相继分化,称为神经期,此时的胚胎称神经胚(neurula)。当胚体发育到出现外鳃、口、尾鳍、心脏跳动和血液循环时,即冲破卵胶膜或卵袋进入水中,孵化成独立生活的幼体——蝌蚪。两栖类的卵从受精到发育成幼体所需的时间,可因时、因地、因水温和种类而不同,通常在水温12℃~23℃的条件下,蛙蟾类约为4~5天,而分布于我国东北地区的极北小鲵则需17~19天。刚出卵胶膜的蝌蚪酷似幼鱼:口的后面有一能分泌粘膜的吸盘,以此吸附在水草上,静止不动,2~3天后吸盘开始退化;有一侧扁的长尾作为运动器官;也有分枝的羽状外鳃执行呼吸机能。外鳃不久就被新发生的鳃盖褶遮蔽起来,并渐趋萎缩而代之以咽部的4对内鳃,鳃盖褶以单个出水孔与体外相通。蛙蟾类蝌蚪的两颌外包黑色的角质喙,构成口的上、下唇附生角质唇齿数行,以此啮食水生植物,唇的边缘另有许多乳突,可能是蝌蚪的味觉感受器。
有些鲵螈类,例如巴尔干半岛地下水中的洞螈、北美的泥螈、中国的山溪鲵和滇池蝾螈(Cy-nopus wolterstorffi)等在性成熟和具有生殖能力时,仍保留着幼体时期的某些特征,这种现象称为幼态成熟(neoteny)。处于幼态时期的动物就能进行生殖的现象,叫作幼体生殖(paedogensis)。分布在墨西哥的虎螈是幼体生殖的典型实例,其幼体名为美西螈(Axolote),栖于水域,用外鳃交换气体,若环境适宜和内分泌生理机能正常,即能进行变态,失去外鳃到陆地上生活(图17-22)。以甲状腺素饲喂虎螈幼体,能激发变态,消失外鳃和尾鳍,成为习惯于离水生活的成体。
在两栖动物的生活史中,蝌蚪必需经过变态(鲵螈类的变态不太明显)才能成为幼蛙或幼鲵(螈)。蝌蚪的变态一般都发生在自由生活了3个月之后,变态期间蝌蚪体内、外出现的一系列变化,实质上是各种器官由适应水栖转变为适应陆生的改造过程。最显著的外形变化是成对附肢的出现、两颌的角质喙及角质唇齿连同表皮一起脱落、蛙蟾类尾部的萎缩消失等。内部器官也有相应变化,当蝌蚪还在以鳃进行呼吸期间,咽部就已经长出了肺芽,并逐渐扩大和形成左、右肺,最终完全代替了鳃。在呼吸器官由鳃转化为肺的过程中,心脏发展成两心房一心室,而血液循环方式也随之由单循环改造成不完全的双循环。完成变态后的幼蛙(蟾)或幼鲵(螈)已能离水登陆营两栖生活,并且演变为吃动物性食物、消化道由螺旋状盘曲转变成粗短的肠管,同时胃、肠的分化也趋于明显(图17-23)。有些两栖动物的蝌蚪期很长,需经过1~2次越冬后才进行变态,因此分布在南美洲乌拉圭及亚马逊河流域的奇蛙,它的蝌蚪体长甚至可以达到250mm而超过成蛙4倍。

第三节 两栖纲分类
现存的两栖纲动物有蚓螈目(Caeciliformes)、蝾螈目(Salamandriformes)和蛙形目(Rani-formes)等3个目,有398属,34科,约4200种。我国产两栖纲动物280余种。
一、蚓螈目(Caeciliformes,无足目Apoda,裸蛇目Gymnophiona)
蚓螈目动物的身体细长,形似蚯蚓,四肢及带骨均退化,无尾或尾极短,是营钻土穴居生活的类型。全身裸露,体表有皮肤褶皱形成的数百条覆瓦状环褶,环褶内有次级环褶及围绕体轴呈环状排列的骨质圆鳞(水生种类无鳞),可用于加固体壁抵抗泥土压力的作用。环褶表面腺体丰富,分泌物能减少水分蒸发,并可有效地降低体表与洞壁的磨擦,加快在洞穴中运动的速度。眼小,大多隐于透明的皮下成眼点状;耳无鼓膜;听神经退化;鼻眼间近颌缘的凹槽内有一能伸缩自如的触突,状如蜗牛的触角,伸长时可达2mm~3mm,敏感而有助于地下钻穴活动。雄性的泄殖腔能翻出体外,用作交配。体内受精,卵生或卵胎生。雌体常抱卵孵化,以皮肤表面的粘液保护卵免致干燥。
蚓螈类是两栖纲中最低等的类群,还保留着一系列原始特征:环褶内的皮下具有来源于真皮的骨质圆鳞;头骨上的膜性硬骨数目多;无荐椎;椎体为双凹型;具长肋骨,但无胸骨;左、右心房间的隔膜发育不完全,动脉圆锥内无纵瓣。
本目共5科、34属、160多种,分布于非洲、美洲和亚洲的热带地区,其中尤以中、南美洲的种类最多。我国仅产一种版纳鱼螈(Ichthyophis bannanicus),属鱼螈科(Ichthyophidae),最早于1974年采自云南省勐腊县,1983年和1985年又先后在广西省十万大山和广东省鼎湖山等地发现了该螈(图17-24)。

二、蝾螈目(Salamandriformes)
蝾螈目又称有尾目(Caudata),形似蜥蜴,四肢细弱,少数种类仅有前肢(鳗螈),终生有发达的尾,尾褶较厚实。皮肤光滑无鳞,表皮角质层薄并定期蜕皮。眼小或隐于皮下(洞螈),水栖种类常缺乏活动性眼睑(大鲵、北美洲的虎螈和泥螈等);无鼓室和鼓膜;少数种类有一对耳旁腺;舌圆或椭圆形,舌端不完全游离,不能外翻摄食;两颌周缘有细齿;有犁骨齿。构成头骨的骨块少,颅侧因无颧骨和方轭骨而边缘不完整。椎体在低等种类(小鲵科、隐鳃鲵科)为双凹型,高等种类则为后凹型;肋骨、胸骨和带骨大多为软骨质;有分离的桡骨、尺骨及尾椎骨。雄性无交配器,体外或体内受精,体外受精种类在水中由两性几乎同时排出卵子和精液,完成授精作用;体内受精种类的雄性先排出由胶质形成的精包(spermatophore),这是泄殖腔内骨盆腺(pelvic gland)和泄殖腔腺(cloaca gland)所分泌的粘性胶质物,连同被包埋在其中的精子共同组成的结构。雌体以泄殖腔壁的外缘将精包纳入其泄殖腔中的受精器内,精包被吞噬细胞破坏而释放出精子,在输卵管内与卵子完成授精作用。绝大多数为卵生,少数卵胎生,是对激流水生环境的一种适应。幼体水栖,有3对羽状外鳃,尾褶较发达;2~3龄时进行变态,但变态不明显,通常以外鳃消失、鳃裂封闭和颈褶形成作为变态结束的标志。
成体栖息于潮湿环境,大多营半水栖生活,也有终生水栖或陆栖的生态类型。以节肢动物、蠕虫、螺类、小鱼、蝌蚪和幼蛙等为食。视觉差,扑食活动主要凭嗅觉或侧线的感觉,求偶时依靠皮肤腺和泄殖腔内的(1~3对)泄殖腔腺分泌物所发出的特殊气味识别同类。除爬行外,主要以四肢后伸贴体和尾部左右摆动的方式在水中游泳前进。再生力强,肢、尾损残后可重新长出再生肢或再生尾。
蝾螈目共9科、60属、约358种,主要分布在北半球,少数渗入热带地区。非洲大陆、南美洲南部和大洋洲无本目动物。我国产约37种,代表科、种有(图17-25):

(一)隐鳃鲵科(Cryptobranchidae)是现存两栖纲动物中体型最大的类群。口大眼小,无眼睑。背部光滑,散有小疣粒,沿体侧有宽厚的纵行肤褶。犁骨齿呈长弧形排列,靠近颌缘并与上颌齿平行。幼体有鳃,成体时消失无迹。椎体双凹型。代表动物为中国大鲵(Andrias davidi-anus),产于我国华南和西南的山地溪流中。体大,呈扁筒形,头部扁宽,眼小,无活动性眼睑。体侧有12~15条肋沟,并有一明显的纵行肤褶,四肢粗壮短小,指、趾间无蹼;背、腹面有许多成对的小疣粒。尾侧扁,有发达的尾褶。栖息于海拔高200m~1600m水质清澈的山间河溪或深潭中,常在多石隙的滩口上下及水流缓慢的洞穴附近活动。昼伏夜出,偶尔登陆,扑食时,常逆水守候在滩口的石堆中,待猎食对象接近时,突然张口咬住,囫囵吞下。11月至翌年3月在潭底石块下冬眠,呈半休眠状态,5~8月为繁殖期,雌鲵一般在雷雨季节的夜间产卵300~500枚,淡黄色的鲵卵在管状胶囊内排列成一长串念珠状。雄鲵常屈体围卵或将卵囊缠在身上,以免敌害吞啮或被水冲去,直至幼体孵化才离开。
中国大鲵是仅产于我国的珍稀动物,古籍中名为鲵、、鱼、啼鱼和孩儿鱼,民间俗称娃娃鱼,也是古老的两栖纲动物,对研究动物的进化和地理分布都有一定意义。体大肉鲜,常是各地盲目滥捕的猎取对象,以致资源曾遭到严重破坏。
(二)小鲵科(Hynobiidae)体较小,全长不超过300mm。有活动性眼睑。犁骨齿成“U”形或排列成左、右两短列。椎体双凹型。皮肤光滑无疣粒,多数种类具颈褶;躯干呈圆柱状,体侧有明显的助沟。体外受精,雌鲵产成对的筒状卵胶囊,卵胶囊呈弧形、环形或螺纹形,一端游离,另一端附着在物体上。本科共7属,约30种,代表种类有:
1.极北小鲵(Salamandrella keyserlingii)全长100~120mm。皮肤光滑,体侧的肋沟往下延伸至腹部;指、趾数均为4枚,无蹼;尾长短于头体长。背面有2条浅褐色或黄棕色纵纹,中间被一深色纵纹隔开。除4月中旬去水沟繁殖产卵外,一般都在低洼地的松土处活动,黄昏和雨后较活跃,夏季炎热时常隐伏于草垛枯木下或距地表30mm~40mm的洞穴内,秋后则聚集在更深的泥土中蛰眠。主要捕食昆虫、蚯蚓和软体动物。分布在我国东北三省、内蒙古;朝鲜、日本和俄罗斯等国。
2.山溪鲵(Batrachuperus pinchonii)全长90mm~220mm。吻端圆,吻棱不显;眼大;上唇褶发达,下唇褶为其所遮盖;舌的两侧游离。指、趾基部无蹼,自指端至肘关节及趾端到跟部覆有极强的棕色角质鞘。体色变异大,背面常有粗大的黑褐色花斑。栖于高山山溪及湖泊石块的树根下、苔藓中或溶雪泉水的碎石间,分布于四川和云南。
(三)蝾螈科(Salamandridae)全长小于200mm。皮肤光滑或有疣瘰,肋沟不显,指4趾5。椎体后四型。有活动性眼睑。犁骨齿呈“∧”形。体内受精,卵单生或连成单行(红瘰疣螈Tylo-totriton verrucosus);大多水中产卵,少数在水源附近的湿土上产卵,成体以水栖为主,也有陆栖种类(疣螈)。本科有16属50余种,广布于北半球温带地区,但在我国却与之不同,所产的6属18种几乎全都分布于秦岭以南地区,这是颇为奇特的现象。代表动物有:
1.肥螈(Pachytriton brevipes)身体粗壮肥硕,背脊正中有一纵形浅沟。头侧无突起的棱嵴。舌大,与口腔底部相连。四肢短粗,趾基有蹼迹。泄殖孔纵列。全身光滑无疣,皮肤富有腺体,能分泌带有特殊的硫黄气味的白色粘液。体浅褐色,背面及尾侧密布黑色圆斑,腹面橘黄色并缀有黑斑。生活在海拔高800m以上水质清凉和流速缓慢的溪流中,也见于水潭和水坑内。白天静匍在石块上或在水底砂石间爬行,遇惊即游向深水区。分布于浙江、江西、广东和广西等省。
2.东方蝾螈(Cynops orientalis)体形较小,全长65mm~85mm。头扁吻钝,吻棱显著,上唇两侧有唇褶。四肢较长而纤弱,指、趾末端尖出,无蹼。尾略短于头体长。背面黑褐色,有分散的小疣粒,腹面朱红色,杂以棕黑色圆斑或条纹。生活于海拔高30m~1000m的山麓或山间静水中,捕食水生昆虫和小形水生动物。3~7月为繁殖季节,卵分批成熟,分期产出,从最早产卵日至产卵结束可延续约一个月,产卵80~100枚,最多可达230余枚。受精卵孵化约20天,幼体大多当年完成变态。广泛分布于江苏、浙江、安徽、江西、福建、湖北、湖南等省。
此外,本目还包括洞螈科(Proteidae)的洞螈和泥螈、鳗螈科(Sirenidae)的鳗螈、两栖鲵科(Amphiumldae)的双趾两栖鲵(Amphiuma means)、无肺螈科(Plethodontidae)的多褶无肺螈(Eurycea multiplicata)、钝口螈科(Ambystomidae)的虎螈和双曲齿螈科(Dicamptodontidae)的大西洋大螈(Dicamptodon ensatus)等。
三、蛙形目(Raniformes)
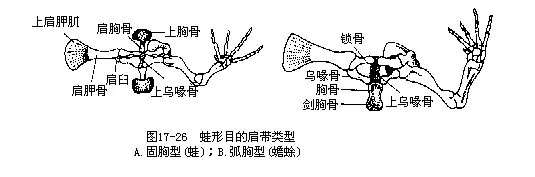
蛙形目是现存两栖纲动物中结构最高等、种类繁多及分布最广的类群。体形短宽,四肢强健,适于跳跃和游泳。成体无尾,故又名无尾目(Anura)。皮肤裸露,内含丰富的粘液腺,有些种类在不同部位集中形成毒腺、腺褶、疣粒等。有活动性眼睑和瞬膜;多数种类具鼓膜。头骨藉方轭骨与上颌骨连接而边缘完整,额骨与顶骨愈合成额顶骨(frontoparietal)。椎体有前凹型、后凹型、变凹型(anomocoelous),或参差型(diplasiocoelous)等不同类型;荐椎后的椎骨合成尾杆骨;一般不具肋骨,胸骨发达。肩带弧胸型(arcifery)或固胸型(firmisterny)(图17-26)。桡骨和尺骨、胫骨和腓骨分别愈合成桡尺骨(radioulna)及胫腓骨(tibiofibula)。变态明显,成体用肺呼吸,营水陆两栖生活。
本目现有20科、303属,约3500种。几遍布热带、亚热带地区,极少数种分布在北极圈内。我国有240多种,代表种类有(图17-27):
(一)盘舌蟾科(Discoglossidae)舌呈圆盘形,舌端无缺刻,舌的四周与口腔粘膜相连,不能伸出口外;仅上颌有齿;雄蟾无声囊。椎体后凹型;第二至第四椎骨具肋骨。肩带弧胸型。我国产铃蟾4种其中有3种局限于云、贵、川和广西境内,仅东方铃蟾(Bombina orientalis)与之相隔间断分布在东北、内蒙古、河北、山东、安徽、江苏等省。
东方铃蟾体形较小,体长约45mm。鼓膜不明显,无耳旁腺。皮肤粗糙,背面灰棕色或绿色,有不规则黑色斑点及许多刺疣,并能分泌毒液。腹面黑色而杂有橘红色或橘黄色的鲜明花斑。受惊遇险时,身体反弓,四肢仰翻,护首挺肚,木然不动,露出醒目的腹部,向进犯者示威,是著名的警戒色范例。如果进犯者对此置若罔闻,张口咬啮,则其口腔粘膜就可能遭到铃蟾背部分泌毒液的强烈刺激。
(二)锄足蟾科(Pelobatidae)瞳孔大多纵立;舌卵圆形,舌端游离而缺刻浅;上颌有齿,通常无下颌齿和犁骨齿。胸侧有胸腺;胁部及股后缘各有一浅色疣粒。趾间无蹼或蹼不发达。肩带弧胸型。椎体变凹型。成体除繁殖产卵期外,很少进入水中。本科共12~14属80余种,分布在欧洲、非洲西北部、亚洲和北美洲,我国产8属约63种,全部生活在海拔较高的南方山区溪流内,较常见的有角蟾类(Magophrys)、髭蟾类(Vibrissaphora)、齿蟾类(Oreolalax)和齿突蟾类(Scutiger)等。
1.宽头大角蟾(Megophrys carinensis)体大,短而肥硕,头体长约100mm,周身均有散在的小圆疣。头大而扁宽,头枕交界处有一横沟;吻棱清晰;上眼睑的外缘有许多锥状疣和1~2个突起的长疣;舌圆宽,舌端略凹。有犁骨齿。前肢粗壮,后肢及趾均短,指、趾末端稍为膨大。背面棕黄色,布满黑斑;两眼间有三角形褐斑,眼后至口角有一深色横纹;腹面有灰纹。栖息在海拔高900m~1500m的山区溪流中,鸣声“啊、啊、啊”。分布于四川、云南、贵州、广西和湖南。
2.崇安髭蟾(Vibrissaphora liui)体长80mm~95mm。头形扁平,吻部宽圆,略突出于下颌;眼球上半部深蓝色,下半部呈棕色;舌端游离,缺刻深;上颌有齿,雄蟾的上唇边缘每侧有1~3枚黑色锥状角刺,雌性在相应部位有橘红色斑点或小黑刺,俗名角怪即由此而来。腹面及体侧满布白色小痣;胯部有白色的月牙斑和股腺。栖息于海拔高800m以上的山间阔叶林带,平时营陆上生活,繁殖期集到山溪上游觅偶交配和产卵。蝌蚪生长缓慢,需经2~3个冬天,才能完成变态。分布于浙江、江西和福建。
(三)蟾蜍科(Bufonidae)体形短粗,背面皮肤上具有稀疏而大小不等的瘰粒。头部有骨质棱嵴;耳旁腺大,其分泌物的干制品即著名中药蟾酥;鼓膜大多明显;瞳孔水平形;舌端游离,无缺刻;无颌齿和犁骨齿。后肢较短。椎体前凹型,无肋骨;肩带弧胸型。陆生性强,昼伏夜出,产卵于长条形的胶质卵带内。本科共13属约335种,分布几乎遍及全球。我国有2属16种,大蟾蜍是习见的种类,此外还有分布在新疆的绿蟾蜍,以及长江以北的花背蟾蜍、青藏高原的西藏蟾蜍和南方地区的黑眶蟾蜍等。

1.花背蟾蜍(Bufo raddei)是分布于长江以北广大地区的优势蟾类。头体长50mm~60mm吻圆,吻棱显著;鼓膜圆形;耳旁腺大而扁。雄蟾背面橄榄黄色,有左、右对称的褐色花斑,分散的灰色疣粒上具有红点,因其斑色艳丽,故名花背蟾蜍。雌蟾背面浅绿色,有酱色花斑及类似异性的疣粒。白天隐伏于洞内,夜间外出捕食。冬季常在鼠洞或农村的地窖中集群蛰眠。
2.黑眶蟾蜍(Bufo melanostictus)头部自吻端沿两侧上眼睑至鼓膜,有突出的黑色骨嵴,嵴后有耳旁腺;鼓膜大而显著。全身除头顶外,布满大小不等的疣粒和圆疣,疣粒上有黑色角刺而显得特别粗糙。背面黑褐色或黄褐色,常饰有不规则的棕色斑纹。黑眶蟾蜍是长江南岸的常见蟾类,夜间外出,行动迟缓,主要食物为作物害虫,有益系数可高达77.88%,在生物防治害虫方面有一定作用。
(四)雨蛙科(Hylidae)小型蛙类。体细瘦,皮肤光滑,无疣粒或肤褶。有上颌齿和犁骨齿。肩带弧胸型。最末2节指骨和趾骨之间各有一间介软骨(intercalary cartilage),指、趾末端膨大成吸盘(digital disc或称指垫pad),并有马蹄形横沟(horse shoe-shaped horizontal groove),适于吸附在挺水植物、农作物和乔灌木的叶上。本科约有35属650种,世界性分布,但以澳大利亚、巴布亚、中美洲、南美洲种类最多,我国产9种,均属于雨蛙属。
日本雨蛙(Hyla japonica)体小,全长不超过45mm。吻短圆而高,吻棱明显,舌端有微缺刻。胸、腹和股腹面有稠密的扁平疣。背部翠绿色或暗绿色,体侧及股的前、后方均无黑斑,腹面白色。雄蛙有单个咽下外声囊,拇指内侧具白色婚垫。生活在平原或山边的稻田附近,鸣声嘹亮而清脆,俗称“梆梆狗”,常与泽蛙、饰纹姬蛙等共栖于同一水域中。除西北和华南地区外,分布几乎遍及全国。与日本雨蛙十分相似的有中国雨蛙(Hyla chinensis),但体侧及股的前后有黑斑,肩部有一个三角形黑斑,第三指的吸盘大于鼓膜,主要分布于我国东南沿海和华南地区。
(五)蛙科(Ranidae)上颌有齿,一般有犁骨齿。鼓膜明显或隐于皮下(湍蛙类、浮蛙类、高山蛙类、倭蛙类),舌端大多具缺刻。椎体参差型,肩带固胸型。本科共40属613种,分布于大洋洲和南极洲以外的各大洲,但以非洲的种类最多。我国产约94种,蛙属几占全部种类的3/4,常见种类有黑斑蛙、金线蛙(Rana plancyi)和日本林蛙(Ranajaponica)等,民间统称为青蛙。其他蛙类有:
1.中国林蛙(Rana chensinensis)体细长,吻端略尖,吻棱发达。背面棕黄色,腹面肉红色,鼓膜区有三角形黑斑;背侧褶明显,脊背及两侧有分散的圆疣或长疣。生活在黄河以北地区,常出没于山溪、水沟和旱生作物的农田间,能分布到海拔高1500m的山上。中国林蛙是我国的传统药用动物,雌蛙整体的干制品即蛤士蟆,干燥的输卵管就是作为滋补药的哈蟆油。
2.泽蛙(Rana limnocharis)体长30mm~40mm。鼓膜不太明显;舌宽,舌端缺刻深;雄蛙有单个咽下外声囊。背部皮肤粗糙,有小圆疣和多行长短不一的肤褶,无背侧褶。体色及斑纹变化甚大,腹面乳黄色。主要生活于稻田、池塘、湖沼附近,捕食活动多在晚间,食物包括蝗虫、蝼蛄、蚱蜢、金龟子等多种农业害虫。分布于黄河以南各省。
3.虎纹蛙(Rana rugulosus)大型蛙类,最大体长超过100mm。头吻尖长,鼓膜大而显著;下颌前侧方有2个骨质齿状突,与上颌的相应凹陷吻合。背面灰棕色或黄绿色,饰有少数深色斑纹;无背侧褶,但有不规则的长短肤褶及散布的小疣粒;腹面白色而平滑。咽胸部有灰黑色斑纹。栖于平原或丘陵地带的山麓附近,常见于水田、池塘和沟渠内,昼伏夜出,机灵敏感,性情凶猛,捕食昆虫的有益系数约75%,有利于农业,但需防范它在养鱼场吞食鱼苗的危害作用。
4.棘胸蛙(Rana spinosa)体大,最长可达140mm。吻圆钝,鼓膜隐约可见。四肢粗壮强健,指、趾末端膨大呈圆球状,趾间全蹼。无背侧褶而有长疣,断续排列成行,并杂有许多小刺疣。背面黑棕色或淡棕色,有浅色斑纹;腹面肉色,布有灰褐色云斑,雄蛙胸部有肉质圆疣,疣上有黑色角刺,棘胸蛙之名即由此而来。生活于山间溪流中,平时隐蔽在石洞里,夜间蹲在潮湿的石块上捕食。
5.海蛙(Rana cancrivora)海蛙分布于东南亚一些热带国家的沿海和我国海南省。栖息在生长着红树林的海湾泥滩上,白天隐藏在红树林根系的隙缝间,傍晚外出到泥滩上觅食小蟹,故又称食蟹蛙。海蛙不但能生活在半咸水或海水等高渗液的环境中,而且还能产卵于海水涨潮时倒灌在岸边的临时性水洼内。孵出的蝌蚪既要有耐盐性,还必需具有承受中午40℃以上高温暴晒的适应能力,这在两栖纲动物中是绝无仅有的。海蛙及其蝌蚪能在海水环境中安然无恙地生活的原因,在于它具有与软骨鱼相似的肾功能和渗透压调节机制,肾脏内肾小球过滤尿素的效率很低,因而造成血液中经常保持着较高浓度的尿素(0.48摩尔浓度),使蛙体内的渗透压高于周围环境。在这种情况下,海蛙的体液不但不会向外渗出,甚至海水中的水分还能透过皮肤不停地渗进体内。
(六)树蛙科(Rhacophoridae)外形及生活习性与雨蛙相似。末端两指、趾节之间有间介软骨,指、趾端明显膨大成吸盘,并有马蹄形横沟。肩带固胸型;椎体参差型。树栖,产卵于卵泡内,蝌蚪生活在静水水域内。全世界约有10属180余种,分布于非洲和亚洲南部。我国产2属43种,分布在秦岭以南各省,常见种类有斑腿树蛙(Polypedates leucomystax),头体长40mm~60mm,雌蛙明显大于异性。吻棱和鼓膜显著;犁骨齿强。指吸盘大,第三指吸盘与鼓膜大小相等,外侧指间无蹼;趾吸盘较指吸盘小,趾间全蹼。体背浅棕色,散有棕黑色斑点,并在前部呈现X形图案,因四肢背面有棕黑色横纹和斑点,以及股后有网纹花斑而名斑腿树蛙。生活在起伏不大的丘陵、山区的水边石缝中。雄蛙的鸣声如击快板,成有节奏的“啪啪……”声。跳跃能力差,行动较迟缓。雌蛙在抱对产卵的同时,还排出胶状物,并以后肢不停地将胶状物搅拌成白色泡沫状卵泡,颜色逐渐由白转黄。雄蛙也在此时排出精液于卵泡上。卵泡大多粘附在草丛中、池塘壁、灌木上或高悬在树枝上。胚胎在卵泡内发育,孵化成蝌蚪后便脱离卵泡或与卵泡一并掉入水中生活。分布于长江以南广大地区。
(七)姬蛙科(Microhylidae)中小型陆栖蛙类。头狭而短,口小,大多数种类无上颌齿和犁骨齿;舌端不分叉。指、趾间无蹼。肩带固胸型。椎体前凹型。蝌蚪的口位于吻端,常缺乏角质颌和唇齿。全世界有60余属约270种,主要分布在非洲、亚洲、大洋洲、美洲的热带地区,我国产4属14种,常见种类有:
北方狭口蛙(Kaloula borealis)体长40mm左右。头和口小,吻部短圆,鼓膜不显。后肢短粗,趾间无蹼。皮肤光滑而厚,背面棕褐色,有不规则的黑色斑点或花纹;腹面肉色。穴居土中,春夏之际在雨后形成的临时性水坑内产卵,分散漂浮,3周内即可完成变态。本蛙是唯一分布在黄河以北的狭口蛙类,与生活于南方的其它狭口蛙存在着明显的间断分布现象。
第四节 两栖类的经济意义
两栖纲动物与人类具有密切关系。绝大多数蛙蟾类生活于农田、耕地、森林和草地,捕食的对象中,常以严重危害作物的蝗虫、蚱蜢、粘虫、稻螟、松毛虫、甲虫、蝽象等所占的比例较高。据统计,平均每只黑斑蛙一天内约捕食70多只昆虫,一只泽蛙捕虫50~270只,两者在全年的食虫数都超过万只,而一只大蟾蜍的灭虫量在3个月中就能达到这个数目。值得一提的是,两栖纲动物捕食的昆虫,常是许多食虫鸟类在白天无法啄食到的害虫或不食的毒蛾等,因而是害虫的重要天敌之一。浙江省于1962—1963年曾对当地22种蛙蟾类进行了食性分析,结果表明身体最小的姬蛙类有益系数最高,达97%,其次是大蟾蜍和雨蛙,为90.14%~93.33%,再次是虎纹蛙、日本林蛙和湍蛙,较低的是树蛙、棘胸蛙、泽蛙和黑斑蛙等。
近年来,利用生物防治有害昆虫,日益受到广大农民的重视。这是因为杀虫农药的耗资大、施用农药会污染环境、农药残毒留存在种子或果实中可以对人体造成慢性毒害,此外害虫对农药的抗药性正在逐年增强,同时抗农药害虫的种类也在不断加多,据联合国卫生组织报道,1908年世界上出现了第一种抗药性害虫,1944年增加到12种,1980年竟高达432种。为此,联合国卫生组织建议世界各国重视和提倡生物防治害虫研究,尽可能避免由于大面积施用农药所带来的弊端。我国福建省莆田县从1974年起在农田中开展了养蛙治虫工作,在每亩稻田内放蛙60只,因蛙大量捕食稻苞虫和螟虫,使防治枯心苗的效果达到91.0%,比每亩稻田施农药的除虫效果88.60%更好。浙江省缙云县农林局病虫观察站曾做过放养黑斑蛙防治稻飞螟和三化螟的试验,同样收到了理想的灭虫效果;由于黑斑蛙还捕食稻纵卷叶螟和稻叶蝉,因而使这两种害虫引起的水稻矮缩病害,也取得明显减少的效果。江西省宜丰县在每亩试验田里放养了400~800只黑斑蛙后,早稻枯心率比喷洒两次农药(甲基1605)的对照田低得多,比未放养蛙的早稻田增产9.2%。由此可见,养蛙治虫的生物防治法不仅是增产节支的有效措施,而且还是防止农药污染环境和作物的理想办法。
保护蛙蟾类除了必须严禁滥捕戮杀外,最重要的是保护它们栖息环境的生态条件,特别注意对于繁殖场地的保护。水体污染和水质恶化是导致两栖纲动物大量死亡的直接原因。农药的毒性对它们及蝌蚪所造成的恶果是严重的;低浓度农药能刺激蝌蚪的肌肉运动,使之易被天敌发现和捕食,高剂量农药则可引起动物迅速死亡。化肥残留物(特别是磷和氮)能改变水体的化学性质,影响卵和蝌蚪的存活。此外,维持两栖纲动物产卵场的水源具有一定深度,也是保证其顺利繁殖的必要条件。

由于蛙肉鲜美可与鸡肉媲美,所以常作为宴席上的珍馔佳肴。然而从保护珍稀动物和有益动物的角度出发,则应大力宣传进行保护,制止为了供给食用而漫无节制地捕杀。原产于美洲的牛蛙(Ranacatesbeiana)已被许多国家移养成为食用蛙种,我国于20世纪50年代末也引进和试验养殖(图17-28)。牛蛙是生活在静水水域中的巨型蛙类,体型肥硕,头体长200mm,重1kg以上,后肢强健粗壮,因其鸣声高亢,犹似牛吼而得名。牛蛙的综合利用价值高,除供食用外,蛙皮还能制革,而其他部分可加工成骨粉和饲料等。近年来,福建、湖南、湖北和陕西等省在加强资源动物保护的同时,还对棘胸蛙和中国大鲵等进行了人工养殖。
很多两栖纲动物可作药用,其中最负盛誉的首推蛤士蟆和蟾酥,市上出售的蛤士蟆是中国林蛙的整体干制品,而其雌性输卵管的干制品即蛤蟆油,富含蛋白质、脂肪、糖、维生素和激素,是我国名贵的强壮健身滋补品。蟾蜍耳旁腺分泌的毒液成分复杂,干制加工后制成的蟾酥,是六神丸、喉症丸、安宫牛黄丸、蟾力苏、清毒解毒丸、梅花点舌丹、痧气丸、蟾酥丸、蟾酥锭等数十种中成药的主要原料。蟾酥在临床上用于急救各种心力衰弱,以及对口腔炎、咽喉肿痛、肿节等有镇痛、消炎、退肿和止血的作用;在口腔外科手术中可用作粘膜表面麻醉剂;对皮肤癌和血液病也有一定疗效。不过,蟾酥含毒,需在医生指导下服用,否则可能引起中毒或甚至危及生命。
两栖类数量多、分布广,它们的卵大而裸露,便于采集,也容易培养和观察,因此是教学和科学研究的良好实验材料,已广泛使用于生物学、生理学、发育生物学、药理学等实验中。我国著名实验生物学家朱冼先生,用带血针刺蟾蜍未受精卵而获得“没有外祖父的癞蛤蟆”的研究成果,就是一个很好的例子。
复习题
1.结合水陆环境的主要差异总结动物有机体从水生过渡到陆生所面临的主要矛盾。
2.试述两栖类对陆生生活的适应表现在哪些方面?其不完善性表现在哪些方面?
3.简要总结两栖纲躯体结构的主要特征。
4.简述两栖纲动物的主要目和科的特征。
5.简要理解文昌鱼与蛙的发育过程有何异同。
6.为什么要保护青蛙?影响青蛙存活的主要因素有哪些?
本文标题:第十七章 两栖纲(Amphibia)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59416.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59416.html
